Overview
- 연구 배경: 감정은 생존에 중요한 역할을 하며, 사회적 행동을 지도하는 데 핵심적인 기능을 수행하는 것으로 알려져 있으나, 그 기전과 신경 기초에 대한 이해는 여전히 제한적임
- 핵심 방법론:
- 감정 이론(다윈, 프로이트, 제임스-랑지, 캐논-바드 등)과 신경과학적 기초(기질, 뇌 회로)를 종합적으로 검토
- 얼굴 표정 해석, 공감, 정서적 시뮬레이션(미러링) 및 정신화(mentalizing)와 같은 사회적 정보 처리 과정 분석
- 주요 기여:
- 감정이 생존 가치가 있는 자극에 반응하며, 사회적 상황에서 협력, 배제, 도덕적 불쾌감 등 다양한 맥락에서 뇌 회로가 공유될 수 있음을 제시
- 감정의 생물학적 기초와 문화적 영향을 통합한 ‘기본 감정’ 이론과 ‘핵심 감정’ 이론의 차이를 체계적으로 설명
- 실험 결과:
- 다윈의 표정 보존 이론을 뒷받침하는 연구(예: 공포와 역겨움 표정의 생리적 적응 기능)와 캐논-바드 이론을 검증한 에피네프린 주사 실험(인지 맥락이 감정 경험에 영향을 미침) 등 주요 사례 제시
- 한계점: 주로 인간과 영장류에 초점을 맞춘 연구로, 다른 생물종에 대한 적용 가능성은 제한적이고, 감정의 문화적 차이에 대한 심층적 분석은 미흡함
📋 목차
대단원 구조
- Theories of Emotion — 감정 이론의 역사적 발전과 현대적 관점 (James-Lange, Cannon-Bard, 기본 감정, 핵심 감정)
- 1.1 Some Characteristics Of Emotions — 감정의 주요 특성과 다윈/프로이트의 초기 이론
- 1.2 James-Lange and Cannon-Bard — James-Lange 이론과 Cannon-Bard 이론의 비교
- 1.3 Papez circuit and the limbic brain — Papez 회로와 변연계 이론
- 1.4 Moral emotions — 도덕 감정의 정의와 인지적 평가 과정
- 1.5 Morality In The Brain — 도덕 판단 시 활성화되는 뇌 영역
- Neural Substrates of Emotion Processing — 감정 처리의 신경 기질 (편도체, 인서라, 안와전두피질, 전대상피질, 복측선조체)
- 2.1 The amygdala: fear and other emotions — 편도체와 공포 조건화
- 2.2 The insula: disgust and interoception — 인서라와 역겨움/내부 감각
- 2.3 Orbitofrontal cortex: contextualized emotions, and emotional feelings — 안와전두피질
- 2.4 Anterior cingulate: response evaluation, autonomic responses, and pain — 전대상피질
- 2.5 Ventral striatum and reward — 복측선조체와 보상
- Reading Faces — 얼굴 표정 인식, 시선 감지, 시뮬레이션 이론
- Reading Minds — 공감, 미러링, 시뮬레이션 이론과 멘탈라이징의 신경 기전
- 4.1 Mind-reading in autism — 자폐 스펙트럼 장애와 마음 이론, 깨진 거울 이론
- 4.2 Neural basis of theory-of-mind — 마음 이론의 신경 기반 (측두극, 내측전두피질, 측두-두정엽 접합부)
- Summary and Key Points of the Chapter — 장 요약 및 핵심 포인트
Chapter 15 The social and emotional brain
Summary
감정은 생존 가치가 있는 자극에 대한 내부 지표로, 사회적 행동을 지도하는 데 필수적이다. 사회적 배제는 신체적 통증과 뇌 회로를 공유하고, 협력을 통한 보상은 보상 회로를 더 강하게 활성화한다. 감정과 별개로 멘탈라이징(perspective taking)에 기반한 비감정적 인지 통제도 사회적 결정에 관여한다.
Emotions act as internal signposts: they guide us how to behave, what to avoid, and what to seek out. Emotions are one way of tagging certain stimuli to ensure that they receive priority treatment and are responded to appropriately. Emotions are linked to stimuli and situations in whi there is an inherent survival value: for instance, fear may be linked to threatening stimuli that require vigilance or withdrawal; disgust may be linked to stimuli relating to contamination; anger may be linked to situations that threaten territory and status; and so on. Although some stimuli may be naturally rewarding (e.g. food, sex) or punishing (e.g. pain), we can learn to assign emotional states to a wide range of novel stimuli su as pop music and fashions, giving rise to extremes from phobias to fetishes. ere is an
almost unlimited flexibility in the range of stimuli that can be linked to emotions even though they may ultimately tap into a narrower repertoire of emotional-related responses (fight, flight, avoidance, etc.) and states (fear, anger, etc.).
Emotions also play a crucial role for guiding social behavior in most social species, including humans and primates. Group living has obvious survival advantages. ere is safety in numbers and cooperation enables the sharing of limited resources. As su, it not surprising that emotions guide social decision making. is apter gives many examples of how the emotional brain is recruited in social situations. For instance, social rejection may share neural circuitry with physical pain, and moral disgust may have something in common with contamination-related disgust. Similarly, the reward circuitry of the brain is activated more if $10 is won by cooperating with another player than if the same amount is obtained without cooperation. It is as if the act of cooperating is a reward in its own right. However, we are not a slave to our emotions. We do not always act on our “gut instincts” and can engage in nonaffective cognitive control to guide behavior. Indeed, thinking about other people’s intentions, desires, and beliefs via perspective taking (or mentalizing) is linked to a rather different brain network to that involved in emotional evaluations.
Key Terms
Emotion
A state associated with stimuli that are rewarding (i.e. that one works to obtain) or punishing (i.e. that one works to avoid). These stimuli often have inherent survival value.
감정(emotion)은 보상성 자극(얻고자 하는 것)이나 처벌성 자극(피하고자 하는 것)과 연관된 상태로, 내재적 생존 가치를 지닌 자극에 대한 적응적 반응을 촉진하는 생리적-심리적 메커니즘이다.
Mentalizing
The process of inferring or attributing mental states to others.
멘탈라이징(mentalizing)은 타인의 정신 상태(의도, 감정, 믿음)를 추론하는 인지적 과정으로, 사회적 상호작용과 행동 예측에 핵심적인 역할을 한다.
Mirroring
The process of sharing the emotions or mental states of others.
미러링(mirroring)은 타인의 감정이나 정신 상태를 자신의 경험과 연결하여 공감하는 심리적 메커니즘으로, 협력, 공감, 사회적 유대감 형성에 핵심적이다.
Mood
An emotional state that is extended over time (e.g. anxiety is a mood and fear is an emotion).
기분(mood)은 시간에 걸쳐 지속되는 감정 상태로(예: 불안=기분, 공포=감정), 이 장에서는 감정 이론, 표정/시선에서의 사회적 정보 추출, 시뮬레이션을 통한 공감, 미러링 또는 멘탈라이징을 통한 타인 정서 이해 과정을 다룬다.
e apter begins by considering various theories of emotion, both historical and contemporary, and then places these theories into the context of the known neuroscientific basis of emotional processing. e apter then considers how social information is extracted from facial expressions and eye gaze. is provides an important introduction to how perceiving emotions (in other people) may result in a simulation of that emotional state in the perceiver (i.e. a sharing of emotion). is idea is taken further in relation to the neural meanisms of empathy. is discussion considers the extent to whi we understand others through a process of simulation of emotional, motor or bodily states (also called mirroring) or inferring mental states (mentalizing, also called theory-of-mind).
Theories of Emotion
Summary
감정은 다면적 현상으로, 이론에 따라 주관적 경험(설문지), 신체 반응(피부 전도도, 표정), 행동 결과(보상/처벌 반응) 등 서로 다른 측면에 초점을 맞추며, 이는 감정 측정의 다양한 방법론을 반영한다.
Emotions are multi-faceted in nature, and the list below captures the key aracteristics. Some theories of emotion have tended to concentrate on some aspects more than others based on the assumption that some features are more core than others. It also means that there are many different ways of measuring emotion depending on whether one concentrates on their subjective nature (e.g. using questionnaires), their bodily responses (e.g. using skin conductance, or recordings of facial expression), or their behavioral consequences (e.g. pressing a lever for a reward).
Some Characteristics Of Emotions
Summary
감정은 보상성/처벌성 자극에 대한 일시적 내재 상태로, 주의 집중, 쾌락적 가치(hedonic value), 신체 반응(심박수, 호르몬), 외부 표현(표정, 자세) 등의 특성을 지닌다. 다윈과 프로이트는 인간 감정이 동물과 연속성을 가진다는 핵심 가정을 공유한다.
- An emotion is a state associated with stimuli that are rewarding (i.e. that one works to obtain) or punishing (i.e. that one works to avoid). ese stimuli oen have inherent survival value.
- Emotions are transient in nature (unlike a mood, whi is where an emotional state becomes extended over time), although the emotional status of stimuli is stored in long-term memory.
- An emotional stimulus directs aention to itself, to enable more detailed evaluation or to prompt a response.
- Emotions have a hedonic value , that is, they are subjectively liked or disliked.
- Emotions have a particular “feeling state” in terms of an internal bodily response (e.g. sweating, heart rate, hormone secretion).
- Emotions elicit particular external motor outcomes in the face and body, whi include emotional expressions . ese may prepare the organism (e.g. for fighting) and send signals to others (e.g. that one intends to fight).
Two early views of emotion came from some well-known figures in science: Charles Darwin (1809–1882) and Sigmund Freud (1856–1939). Although their approaes are very different from ea other, they share the fundamental assumption that human emotions possess continuity with their animal counterparts.
Key Terms
Expression
External motor outcomes in the face and body associated with emotional states.
다윈(1872)은 감정 표현이 종 간 공유되는 본능적 특성으로, 공포 표정은 시야 확대를 통해 위험 감지를, 역겨움 표정은 오염 물질 회피를 촉진하는 적응적 기능을 갖는다고 주장했다. 현대 연구에서는 Ekman의 기본 감정 이론이 다윈의 표정 연구를 계승하고 있다.
In 1872, Charles Darwin published “e Expression of the Emotions in Man and Animals” (Darwin, 1872/1965). For mu of this work Darwin was concerned with documenting the outward manifestations of emotions expressions—in whi animals produce facial and bodily gestures that aracterize a particular emotion su as fear, anger, or happiness. Darwin noted how many expressions are conserved across species; anger involves a direct gaze with mouth opened and teeth visible, and so on. He claimed that su expressions are innate “that is, have not been learnt by the individual.” Moreover, su expressions enable one animal to interpret the emotional state of another animal; for example, whether an animal is likely to aa, or is likely to welcome a sexual advance. Darwin’s contribution was to provide preliminary evidence as to how emotions may be conserved across species. His reliance on expressions resonates with some contemporary approaes, su as Ekman’s aempts to define “basic” emotions from crosscultural comparisons of facial expressions (Ekman et al., 1972). More recent resear has elucidated the functional origins of some of these
expressions. For instance, a posed fear expression increases the visual field and nasal volume and leads to faster eye movements (adaptive for detecting danger), whereas a disgust expression has an opposite effective (adaptive for avoiding contaminants) (Susskind et al., 2008).

📊 그림 설명
다윈이 제시한 종 간 감정 표현의 보존을 보여주는 그림이다. 인간과 동물(예: 개, 고양이)의 분노, 공포 등 감정 표현이 유사한 패턴을 공유하고 있음을 시각적으로 비교한다. 이는 감정 표현이 진화적으로 보존된 본능적 특성임을 뒷받침하는 핵심 증거로 활용된다.
Darwin argued that many emotional expressions have been conserved by evolution. From Ward, 2012, p. 73.
For Freud, our minds could be divided into three different kinds of meanisms: the id, the ego, and the super-ego (e.g. Freud, 1920/2010). e id was concerned with representing our “primitive” urges that connect us to nonhuman ancestry including our basic emotional needs for sex, food, warmth, and so on. e id was concerned with unconscious motivations, but these ideas would sometimes be accessible via the ego (the conscious mind), and perhaps conflict with our super-ego (our cultural norms and our aspirations). Freud’s basic idea that emotions are an unconscious bias on our behavior is very mu relevant to current thinking (Tamieo & De Gelder, 2010). Freud’s other enduring influence is the notion that many psyiatric disorders (su as anxiety) can be understood as emotional disturbances (Le Doux, 1996). Although some of the general approa is recognizable today, the specific details of Freud’s theory no longer have contemporary currency (e.g. ideas relating to ildhood sexual fantasies).
Key Terms
James-Lange theory
The self-perception of bodily changes produces emotional experience (e.g. one is sad because one cries).
James-Lange 이론은 신체적 변화의 자가 인식이 감정 경험을 생성한다고 주장한다(예: 울기 때문에 슬프다). 감정은 신체 반응(심박수, 근육 긴장)의 인식을 통해 형성된다.
James-Lange and Cannon-Bard
Summary
Schachter와 Singer(1962)의 에피네프린 주사 실험은 신체 변화만으로는 감정이 유발되지 않지만, 인지적 맥락과 결합하면 감정 경험이 강화됨을 보여주었다. 이는 James-Lange 이론의 한계를 드러내며, Cannon-Bard 이론은 감정 인식과 신체 반응이 별도의 신경 경로를 통해 동시 발생한다고 설명한다.
According to the James–Lange theory of emotion, it is the self-perception of bodily anges that produces emotional experience (James, 1884). us, anges in bodily state occur before the emotional experience rather than the other way around. We feel sad because we cry, rather than we cry because we feel sad. is perspective seems somewhat radical compared with the contemporary point of view. For instance, it raises the question of what type of processing leads to the ange in bodily states and whether this early process could itself be construed as a part of the emotion.
Changes in the body are mediated by the autonomic nervous system (ANS), a set of nerves located in the body that controls activity of the internal organs (the soma). ere is good empirical evidence to suggest that anges in bodily states, in themselves, are not sufficient to produce an emotion. Sacter and Singer (1962) injected participants with epinephrine (also termed adrenaline), a drug that induces autonomic anges su as to heart rate. ey found that the presence of the drug by itself did not lead to self-reported experiences of emotion, contrary to the James–Lange theory. However, in the presence of an appropriate cognitive seing (e.g. an angry or happy man enters the room), the participants did self-report an emotion. A cognitive seing, without epinephrine, produced less intense emotional
ratings. is study suggests that bodily experiences do not create emotions (contrary to James–Lange), but they can enhance conscious emotional experiences.
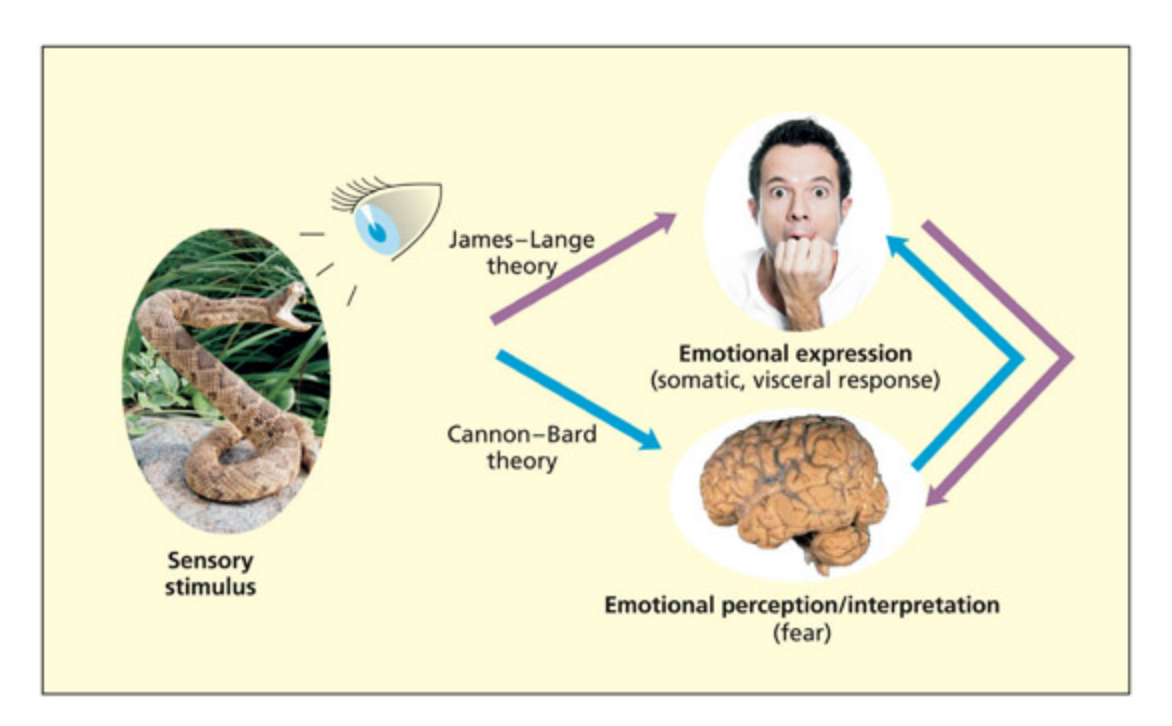
📊 그림 설명
James-Lange 이론과 Cannon-Bard 이론의 감정 처리 경로를 비교하는 다이어그램이다. James-Lange 이론에서는 신체 반응이 먼저 발생하고 그 인식이 감정 경험을 생성하는 반면, Cannon-Bard 이론에서는 감정의 인식/해석이 먼저 발생하고 신체 반응이 뒤따른다. 두 이론의 핵심적인 차이를 화살표 흐름으로 명확히 대비하고 있다.
According to the James–Lange theory, bodily reactions occur first and emotional processing occurs aer (as the perception/interpretation of those reactions). According to the Cannon–Bard theory, the emotional perception/interpretation occurs first and the bodily reaction occurs aer. From Ward, 2012, p. 75.
Key Terms
Cannon-Bard theory
Theory centered on the hypothalamus’ role in emotions in which bodily responses occur after the emotion itself.
Cannon-Bard 이론은 시상하부(hypothalamus)가 감정의 핵심 구조이며, 감정이 먼저 발생한 후 신체 반응이 따른다고 주장한다. 이는 신체 반응이 감정에 선행한다는 James-Lange 이론과 대비된다.
Papez circuit
A limbic-based circuit that was once thought to constitute a largely undifferentiated “emotional” brain.
Papez 회로는 변연계(limbic system) 기반의 신경 회로로, 과거에는 비분화된 감정 처리 기능으로 간주되었으나, 현대 연구에서는 구성 영역들이 다양한 기능적 구분을 가짐이 밝혀졌다.
Basic emotions
Different categories of emotions assumed to be independent of culture and with their own biological basis (in terms of evolution and neural substrate).
기본 감정(basic emotions)은 문화와 무관하게 독립적으로 존재하며, 고유한 진화적/신경적 기반을 가진다. Damasio의 체감 표지 가설은 신체 반응이 무의식적으로 의사결정을 지도한다고 주장하며, Cannon-Bard 이론은 시상하부가 감정을 평가한 후 자율 신경계와 대뇌피질에 동시 신호를 전달한다고 설명한다.
ere are several contemporary theories that bear similarity to the James-Lange theory, most notably Damasio’s (1994) suggestion that bodily responses linked to emotions guide decision making. is is the Somatic Marker Hypothesis discussed in more detail in Chapter 14. Although James-Lange theory states that these bodily responses must be consciously perceived, Damasio (1994) takes the different view that they are unconscious modifiers of behavior.
e Cannon-Bard theory of emotions that emerged in the 1920s argued that bodily feedba could not account for the differences between the emotions (Cannon, 1927). According to this view, the emotions could be accounted for solely within the brain and that bodily responses occur after the emotion itself. e Cannon-Bard theory was inspired by neurobiology. Earlier resear had noted that animals still exhibit emotional expressions (e.g. of rage) aer removal of the cortex. is was considered surprising given that it was known that cortical motor regions are needed to initiate most other movements (Frits & Hitzig, 1870). In a series of lesion studies, Cannon and Bard concluded that the hypothalamus is the centerpiece of emotions. ey believed that the hypothalamus received and evaluated sensory inputs in terms of emotional content, and then sent signals to the
autonomic system (to induce the bodily feelings discussed by James) and to the cortex (giving rise to conscious experiences of emotion).
시험 팁
James-Lange vs Cannon-Bard를 구분하는 핵심: James-Lange = “Body First”(신체 반응이 먼저, 그 인식이 감정), Cannon-Bard = “Brain First”(뇌가 먼저 평가, 신체 반응은 동시/이후). Schachter-Singer 실험은 둘 다 부분적으로 맞다는 것을 보여준다. 에피네프린 주사만으로는 감정이 생기지 않지만(James-Lange 반박), 인지적 맥락과 결합하면 감정이 강화된다(신체 반응의 역할 인정).
Papez circuit and the limbic brain
Summary
Papez 회로와 MacLean의 변연계 이론은 단일 감정 회로를 제안했으나, 해마와 시상하부가 감정에 국한되지 않고 공포/혐오 등이 서로 다른 신경 기반을 가진다는 점에서 더 이상 지지받지 않는다. 현대 이론은 Ekman의 기본 감정(6가지 범문화적 감정)과 Feldman-Barrett의 핵심 감정(core affect; 쾌락/불쾌 + 흥분도 두 축)으로 나뉘며, Rolls의 구성주의적 접근도 감정이 보상/처벌의 조합으로 구성된다고 본다.
Papez (1937) drew upon the work of Cannon-Bard in arguing that the hypothalamus was a key part of emotional processing, but extended this into a circuit of other regions that included the regions of the cingulate cortex, hippocampus, hypothalamus and anterior nucleus of the thalamus. Papez argued that the feeling of emotions originated in the sub-cortical Papez circuit whi was hypothesized to be involved in bodily regulation. A second circuit, involving the cortex, was assumed to involve a deliberative analysis that retrieved memory associations about the stimulus. e work of MacLean (1949) extended this idea to incorporate regions su as the amygdala and orbitofrontal cortex, whi he termed the “Limbic Brain.” e different regions were hypothesized to work together to produce an integrated “emotional brain.”
ere are a number of reasons why these earlier neurobiological views are no longer endorsed by contemporary cognitive neuroscience. First, some of the key regions of the Papez circuit can no longer be considered to carry out functions that relate primarily to the emotions. For example, the role of the hippocampus in memory was not appreciated until the 1950s (Scoville & Milner, 1957), and the hypothalamus is not a central nexus of emotions although it does regulate bodily homeostasis. Second, contemporary resear places greater emphasis on different types of emotion (e.g. fear versus disgust) having different neural substrates.
Contemporary views of emotion: categories, dimensions, and appraisals
e dominant alternative view to that of an undifferentiated “limbic brain” in the contemporary literature is to postulate different categories of emotion (e.g. fear, anger, disgust). However, within this broad framework there are very different views as to where su categories emerge from. In one approa, called the basic emotions approa (Ekman, 1992), there are postulated to be small number of distinct emotions that have been shaped by different evolutionary demands, are linked to distinct neural substrates in the brain, and have specific facial expressions that do not vary across cultures. e alternative contemporary approa is to argue that different categories of emotions are constructed from different kinds of core processes su as autonomic responses, approa/avoidance reactions, and other ongoing cognitions (e.g. beliefs, appraisals). ese theories do not dismiss the notion of qualitatively different types of emotion (su as disgust or fear). ey make the claim that these categories are not “natural kinds,” but rather represent different points within a broader space of emotional experience (Feldman Barre, 2006).

📊 그림 설명
Paul Ekman이 다양한 문화권에서 수집한 6가지 기본 감정(슬픔, 기쁨, 역겨움, 놀람, 분노, 공포)의 얼굴 표정 사진이다. 각 감정에 대응하는 보편적 표정 패턴을 보여주며, 이는 기본 감정이 문화를 초월하여 공유되는 생물학적 특성임을 시사한다.
Paul Ekman tested a wide range of different cultures and concluded that there are six basic types of emotion expressed in faces: sad, happy, disgust, surprise, anger and fear.
© Paul Ekman. Reproduced with permission.
One of the most influential ethnographic studies of the emotions concluded that there are six basic emotions that are independent of culture (Ekman & Friesen, 1976; Ekman et al., 1972). ese are: happy, sad, disgust, anger, fear and surprise. ese studies were based on comparisons of the way that facial expressions are categorized and posed across diverse cultures. Ekman (1992) considers other aracteristics for classifying an emotion as “basic” aside from universal facial expressions, su as ea
emotion having its own specific neural basis; ea emotion having evolved to deal with different survival problems; and occurring automatically. is approa to emotions has been very influential within cognitive neuroscience. However, it encounters various problems. It doesn’t appear to be the case that ea basic emotion has its own unique set of brain regions or networks (although some degree of specialization is found for some categories). Some emotions may also appear to “basic” in some ways but not others: for instance, the emotion of love reflects a clear evolutionary adaptation (for nurturing) and has some specialized neural circuitry, but isn’t linked to a facial expression.
Not all contemporary theories endorse the idea of distinct emotional categories. Two will be considered here, namely the theories of Feldman-Barret (2006) and Rolls (2005). e theory of Feldman-Barre and colleagues (Barre & Wager, 2006; Feldman Barre, 2006; Lindquist & Barre, 2012) assumes that all emotions tap into a system termed core affect that is organized along two dimensions: pleasant-unpleasant and high/low arousal. e laer is also termed activation. Evidence that emotional experience can be classified along these two dimensions comes from studies employing factor analysis of current mood ratings (Yik et al., 1999). In biological terms, this is linked to bodily feelings of emotion and linked to limbic structures su as medial temporal lobes, cingulate and orbitofrontal cortex (Lindquist & Barre, 2012). is eoes the older ideas of Papez and Maclean. e novel aspect of the model is the idea that categories of emotion are constructed (and can be differentiated from ea other) because they tap the core affect system in somewhat different ways and because they are linked to certain kinds of information processed outside of the core affect system, including executive control (for regulating and appraising emotions), language (for categorizing and labeling), theory-of-mind (for conceptualizing emotions in terms of other agents), and so on.
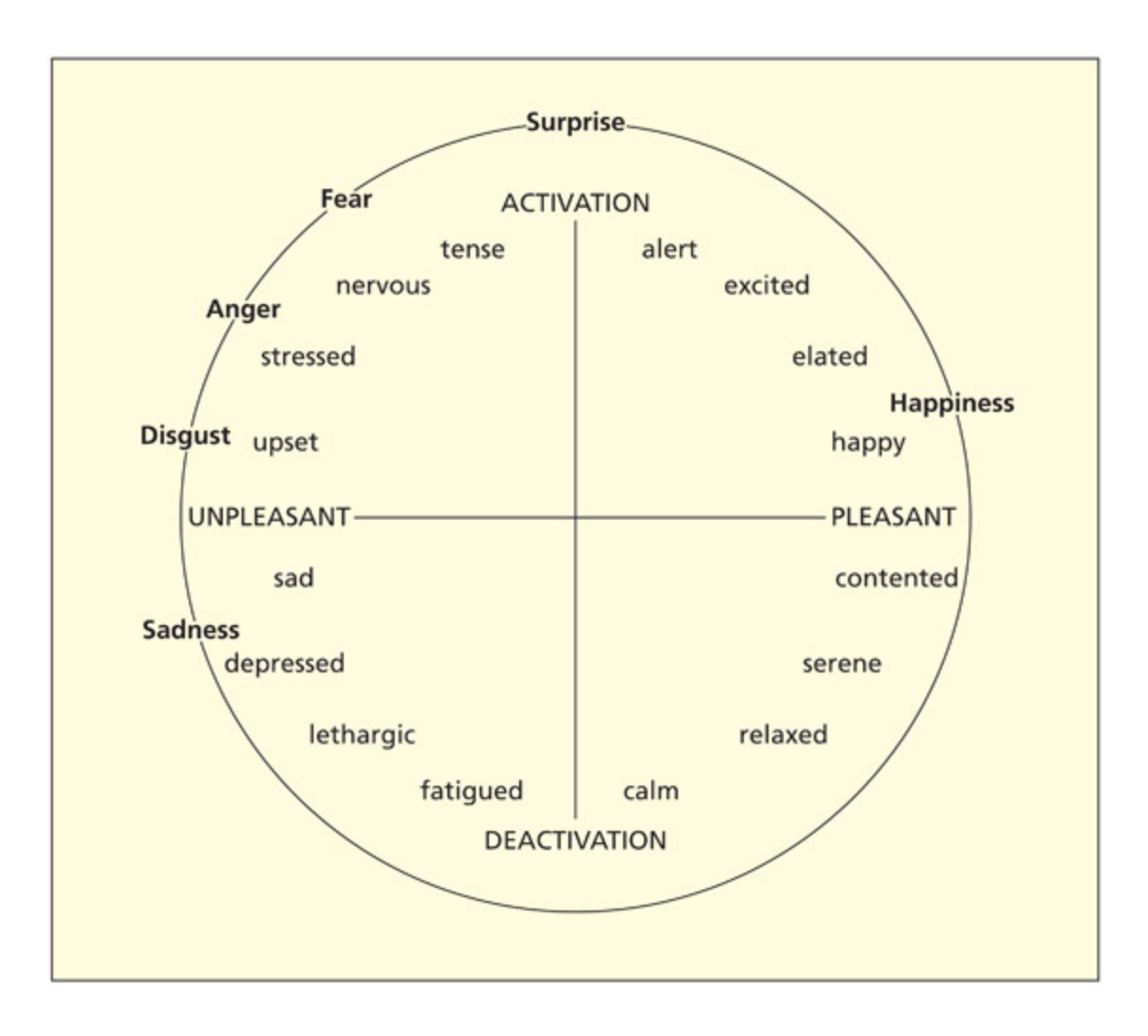
📊 그림 설명
Feldman-Barrett의 핵심 감정(core affect) 모델을 2차원 공간으로 시각화한 그림이다. 가로축은 쾌락-불쾌(pleasantness), 세로축은 각성도(arousal/activation)를 나타내며, 다양한 감정 범주(예: 공포, 기쁨, 슬픔)가 이 2차원 공간의 서로 다른 지점에 위치한다. 이 모델에서는 개별 감정이 특별한 지위를 갖지 않으며, 언어, 기억, 인식 등과의 연합을 통해 구성된다.
In the model of Feldman-Barre, all emotions (and mood) involve a “core affect” system that is organized along two dimensions corresponding to pleasantness and arousal (/activation). Different categories of emotion are points in that space (and linked to associated cognitions—language, memory, perception, theory-of-mind) but are not afforded a special status.
From Russell and Feldman-Barre, 1999. Journal of Personality and Social Psychology.
Finally, the theory of Rolls (2005) also argues for a constructionist approa to emotions, without relying on the notion of a core set of basic emotions. However, his account is different in detail to that of Feldman-Barre (2006). Rather than the notion of “core affect” (with dimensions of arousal and pleasantness), the central part of Rolls’ theory is concerned with the dimensions of reward and punishment, their presence/absence and intensity. Different types of emotion emerge by considering whether a
reward or punishment is applied (e.g. pleasure compared with fear), whether a reward is taken away (e.g. anger) or a punishment is taken away (e.g. relief). ese may occur in combination: for example guilt may be a combination of reward and punishment learning. In addition, the context in whi an emotional stimulus appears is important and an integral part of the constructed emotion. For example, whether the stimulus is social or not (i.e. related to other people) may determine whether the emotion feels like love, anger, jealousy (emotions implying another agent) versus enjoyment, frustration or sadness (emotions that need not imply another agent). Indeed the eliciting stimulus is considered part of the emotional state, so love for one person may be different to love to another person just because the individual is different.
Key Terms
Moral emotions
Emotions that are related to the behavior of oneself (in relation to others) or the behavior of others (in relation to oneself or others).
도덕 감정은 자신/타인의 행동이 규범적 기준에 비추어 평가될 때 발생하며(Haidt, 2003), 인지 평가(appraisal)에 따라 동일한 감정이 부끄러움 또는 죄책감으로 해석될 수 있다. 이는 진화적으로 오래된 감정 기전과 자기/타인 행동에 대한 반성적 능력의 결합에 기반하며, 자랑, 부끄러움, 감사는 인간 고유의 감정일 수 있다.
It is important to note that advocates of the “basic emotions” approa do not deny that some emotions are constructed, but they differ from theories su as those of Feldman-Barre (2006) and Rolls (2005) that assume that all emotions are constructed. One possibility within the basic emotion approa is to consider some emotions as being comprised of two or more basic emotions: for example, joy + fear = guilt, and fear + surprise = alarm (Plutik, 1980). Another possibility is that some emotions are constructed from a basic emotion(s) plus a nonemotional cognitive appraisal. An appraisal of an emotion involves an evaluation of both the content (e.g.
negative feeling) and the context. So a similar feeling could be appraised as either shame or guilt depending on whether it is contextualized relative to the self (shame) or other (guilt). Haidt (2003) has used the term moral emotions to refer to emotions that are related to the behavior of oneself (in relation to others) or the behavior of others (in relation to oneself or others). It implies the existence of some normative benmark with whi to evaluate our actions. ese norms could be a product of both innate meanisms (e.g. an instinctive desire not to harm others) and culturally accepted norms (e.g. law and religion). In this view, the existence of moral emotions depends on an evolutionarily older set of emotional processes together with an evolutionarily newer ability to reflect on the behavior of self and others. Along these lines, Smith and Lazarus (1990) argue that pride, shame, and gratitude might be uniquely human emotions. Darwin (1872) also believed that blushing (linked to shame or embarrassment) might be a uniquely human expression.
Evaluation
Summary
감정은 진화적 적응 가치를 지니며 종 간 보존되어 있고, 의식적/무의식적 과정과 자율 신경계를 통한 뇌-몸 상호작용을 포함하는 다면적 현상이다. 도덕 감정(예: 죄책감, 자랑)은 감정적 메커니즘과 인지적 평가(appraisal)의 조합으로 구성된다. 현대 이론에서는 감정 범주가 자연적 차이를 반영하는지, 보상/처벌/흥분/평가 등 기초 과정의 조합인지에 대한 논쟁이 지속되고 있다.
Although there are many different theories of emotion (some in vogue, some rejected), there are a core set of ideas concerning emotions that have stood the test of time. is includes the idea that emotions have an evolved adaptive value, and this is largely conserved across species. It also includes the notion that emotions are multi-faceted: they contain both conscious (at least in humans) and unconscious processes; they involve the interplay of brain and body via the autonomic system (although emotions cannot be reduced to bodily sensations); and that (at least in humans) some emotions are constructed from both affective meanisms and cognitive ones (e.g. appraisal). A good example of the laer is the so-called moral emotions (e.g. guilt, pride). Contemporary theories emphasize categorical distinctions between emotions (su as anger, fear sadness) but differ with regards to whether these categories represent natural kinds (i.e. innately specified categorical differences, as in the basic emotion approa) or are themselves
constructed from different combinations of building blos of other kinds of core processes (e.g. reward/punishment, pleasure, arousal, appraisals). is idea will be returned to again in the next section.
Morality In The Brain
Summary
Moll 등(2002)의 fMRI 연구에서 도덕 위반 이미지는 일반 정서 자극 대비 안와전두피질, 내측전두피질, STS를 추가적으로 활성화했다. 안와전두피질 손상 환자는 공감, 죄책감 등 도덕 감정이 손상되어, 도덕 딜레마에서 감정보다 논리적 결정을 우선시하는 경향을 보인다.

📊 그림 설명
도덕 위반 장면(예: 가정 폭력)을 관찰할 때 활성화되는 뇌 영역을 보여주는 fMRI 결과이다. 도덕 위반 이미지는 일반적인 정서 자극과 비교하여 안와전두피질, 내측전두피질, 상측두구(STS)에서 추가적인 활성화를 유발하며, 이는 도덕 감정이 일반 감정과 구분되는 신경 기반을 가짐을 시사한다.
What regions of the brain are activated when viewing (or thinking about) scenes involving moral transgressions, su as domestic violence? Is it the same paern found when viewing other emotional stimuli that do not involve a transgression?
Moral judgments involve an evaluation of actions and intentions (either our own or that of others) against some standard of acceptable behavior. Moral emotions occur when we compare behavior against those standards (Haidt, 2003). For instance, if our own actions exceed our standards then we may feel pride, but if they fall below those standards we may feel shame, guilt, or embarrassment. If other people’s behavior falls below standard then we may feel anger or disgust. e question of where the moral standards come from is an interesting one. It is likely to derive from a core set of instincts around
love for one’s family, need for affiliation, empathy, and fairness (including retribution against unfairness).Cultural norms, including religion and the law, tend to uphold this (can you think of a successful religion that does not prea love for one’s family and in-group?). But they may extend moral norms in more idiosyncratic ways (e.g. what to eat and wear).
ere is evidence consistent with the view that processing of moral emotions involves brain structures involved in both emotion and in cognitive appraisal. Moll et al. (2002) presented pictures of three kinds of emotional scenes to participants undergoing fMRI: images of moral violations (e.g. images of physical assaults, abandoned ildren), images of aversive scenes (e.g. dangerous animal) and pleasant images. ese were mated for their self-reported arousal. e moral-violation and aversive images were mated in terms of how negatively they were judged, but the moral violation images were judged as more morally unacceptable than the other affective stimuli. All affective stimuli (relative to a neutral set of images) tended to activate regions linked to emotional processing su as the amygdala and insula, but moral emotions (relative to other affective stimuli) additionally activated regions su as the orbitofrontal cortex, the medial prefrontal cortex and the right posterior superior temporal sulcus (STS). e medial prefrontal cortex and right posterior STS have been linked to theory-of-mind (Amodio & Frith, 2006; Saxe, 2006), whereas the orbitofrontal cortex is implicated in the regulation of social behavior. Similar results were obtained for the moral emotions of embarrassment (Berthoz et al., 2002) and guilt (Takahashi et al., 2004) elicited by reading verbal narratives; for example, “I le the restaurant without paying” (guilt) and “I mistook a stranger for my friend” (embarrassment).
Patients with acquired lesions to the orbitofrontal (and ventromedial prefrontal) cortex oen display poor social functioning (see Chapter 14 for more discussion). ese patients are judged by family members to exhibit low levels of empathy, embarrassment and guilt (Koenigs et al.,
2007). at is, their impairments extend to the moral emotions. When given certain moral dilemmas they tend to perform atypically. For instance, if asked whether they would be willing to push one person under a train to save the lives of five people they are inclined to agree with this course of action (Koenigs et al., 2007). e explanation for this is that there are two conflicting answers in this dilemma. ere is a numerically logical answer that killing one life is beer than killing five lives. ere is also a more emotionally loaded proposition, namely that it would be wrong to push someone under a train. In patients with orbitofrontal lesions, logic may win when pied against a moral emotion.
임상 사례
유명한 트롤리 딜레마(trolley problem)에서 안와전두피질 손상 환자는 “한 사람을 밀어 다섯 명을 구하겠다”고 더 쉽게 답한다. 이는 도덕적 혐오감이라는 감정적 브레이크가 손상되어 순수한 수치적 논리만 남기 때문이다. 이 사례는 도덕 판단이 순수한 이성이 아니라 감정과 인지의 상호작용임을 보여주며, Damasio의 체성 표지 가설(Somatic Marker Hypothesis)과도 연결된다.
Key Terms
Amygdala
Part of the limbic system, implicated in learning the emotional value of stimuli (e.g. in fear conditioning).
편도체(amygdala)는 변연계의 일부로, 두려움 조건화 등 자극의 정서적 가치 학습에 핵심적이다.
Kluver-Bucy syndrome
In monkeys after bilateral amygdala and temporal lesions, an unusual tameness and emotional blunting; a tendency to examine objects with the mouth; and dietary changes.
Kluver-Bucy 증후군은 양측 편도체와 측두엽 손상으로 이상적 온순함, 정서적 둔화, 구강 탐색 경향, 식이 변화가 나타나며, 편도체가 정서적 자극의 학습과 사회적 행동 조절에 핵심적임을 보여준다.
Neural Substrates of Emotion Processing
Summary
감정 처리에 관여하는 주요 뇌 영역은 사회적/비사회적 자극 모두에 적용되며, 기본 감정이 서로 다른 신경 기질을 가지는지에 대한 증거를 통해 감정 이론 간 논쟁을 검증한다.
is section introduces many of the key brain regions involved in emotional processing and considers their possible functions. e section will show how the same brain networks are used to process both social stimuli (our perceptions and interactions with others) as well as nonsocial stimuli with affective properties (su as snakes, food, electric shos). Another aim of the current section is to use this evidence to adjudicate between various theories in the field: for instance, to determine whether there are basic emotions with distinct neural substrates.
The amygdala: fear and other emotions
Summary
편도체는 측두엽 전방에 위치하며, 두려움 조건화(fear conditioning)에서 중립 자극과 충격의 연합 학습 및 저장에 핵심적이다. 기저측 핵(basolateral nucleus) 손상은 학습과 기억 형성을 방해하며, 편도체 손상 동물은 기존의 공포 자극에는 반응하지만 새로운 연합 학습에 실패한다.
e amygdala (from the Latin word for almond) is a small mass of gray maer that lies buried in the tip of the le and right temporal lobes. It lies to the front of the hippocampus and, like the hippocampus, is believed to be important for memory—particularly for the emotional content of memories (Riardson et al., 2004) and for learning whether a particular stimulus/response is rewarded or punished (Gaffan, 1992). In monkeys, bilateral lesions of the amygdala have been observed to produce a complex array of behaviors that have been termed the Kluver–Bucy syndrome (Kluver & Bucy, 1939; Weiskrantz, 1956). ese behaviors include an unusual tameness and emotional blunting; a tendency to examine objects with the mouth; and dietary anges. is is explained in terms of objects losing their learned emotional value. e monkeys typically also lose their social standing (Rosvold et al., 1954).
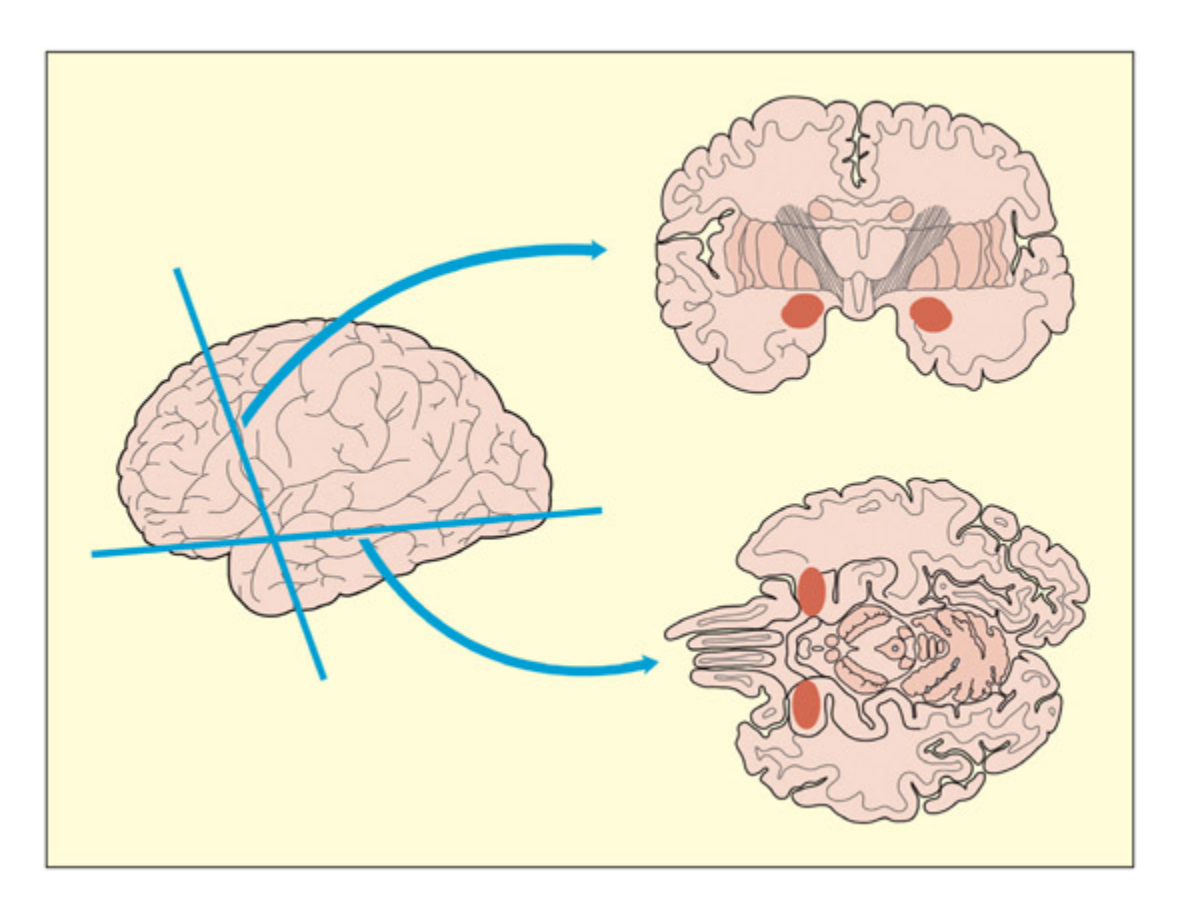
📊 그림 설명
편도체(amygdala)의 해부학적 위치를 보여주는 뇌 단면도이다. 편도체는 좌우 측두엽 전방 내측에 양측으로 매몰되어 있으며, 해마(hippocampus) 앞에 위치한다. 이 영역은 정서적 학습, 특히 두려움 조건화에서 핵심적인 역할을 수행한다.
e amygdala is buried, bilaterally, in the anterior portion of the temporal lobes.
e role of the amygdala in fear conditioning is well established (Le Doux, 1996; Phelps, 2006). If a stimulus that does not normally elicit a fear response, su as an auditory tone (unconditioned stimulus, CS–), is paired with a stimulus that does normally evoke a fear response (termed conditioned response), su as an electric sho, then the tone will come to elicit a fear response by itself (it becomes a conditioned stimulus, CS+). If the amygdala is lesioned in mice (specifically the basolateral nucleus of the amygdala) then the animal does not show this learning, and if the lesion is performed aer the animal has been trained then this learned association is lost (Phillips & Ledoux, 1992). at is, the amygdala is important for both learning and storing the conditioned fear response (although for a different view see Cahill et al., 1999). Single cell recordings suggest that different cells within the amygdala could be involved in learning versus storage of the association (Repa et al., 2001). Animals with lesions to the amygdala still
show a fear response to normal fear-evoking stimuli (su as shos) whi suggests that its role is in learning and storing the emotional status of stimuli that are initially emotionally neutral.
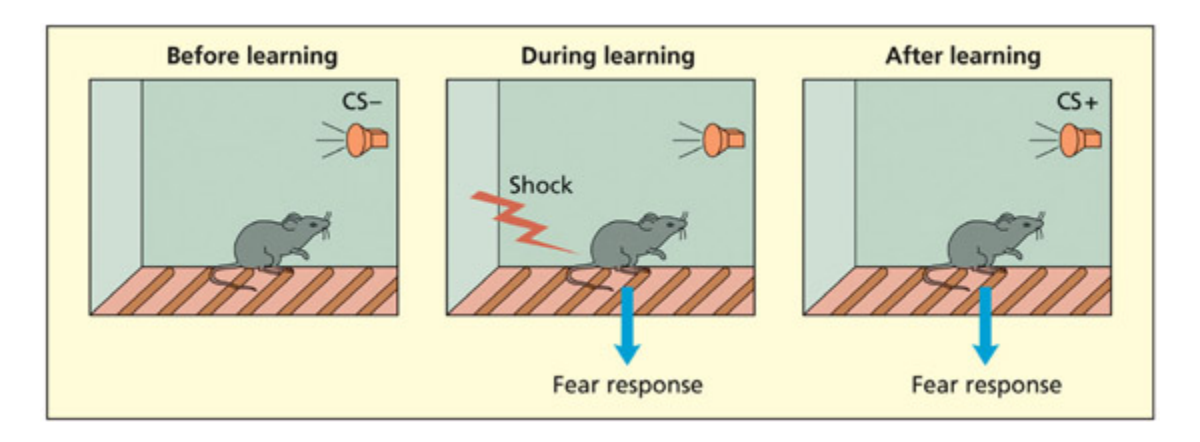
📊 그림 설명
두려움 조건화(fear conditioning)의 기본 절차를 도식화한 그림이다. 처음에는 중립적인 자극(CS-, 예: 소리)이 전기 충격과 함께 반복 제시되며, 충분한 반복 후 소리만으로도 공포 반응을 유발하게 된다(CS+). 이 과정에서 편도체가 자극-감정 연합의 학습과 저장에 핵심적인 역할을 담당한다.
e basic procedure in fear conditioning involves presenting an initially neutral stimulus (the CS–, e.g. a tone) with a sho. Aer sufficient pairings, the stimulus will elicit a fear response without an accompanying sho (it has become a CS+).
From Ward, 2012, p. 83.
Key Terms
Skin conductance response (SCR)
Changes in electrical conductivity on a person’s skin, triggered by certain stimuli (e.g. emotional or familiar stimuli).
SCR(피부 전도 반응)은 자율 신경계 흥분을 측정하는 지표이다. 편도체 손상 환자는 조건화된 공포 반응(SCR)을 보이지 못하지만 연관성을 언어적으로 학습할 수 있고, 해마 손상 환자는 반대 패턴을 보여 두 구조가 병렬적으로 연합을 저장함을 시사한다. LeDoux(1996)는 편도체가 하위피질 경로를 통해 위협에 빠르게 반응하며, 편도체는 단순한 “공포 중심”이 아니라 자율 신경계, 시각 피질 등과 연결된 네트워크의 핵심 허브이다.
In humans, a comparison of learned fear responses to a sho (CS+) with neutral stimuli (CS–) reveals amygdala activation during fMRI that correlated with the degree of conditioned response, in this instance a skin conductance response (LaBar et al., 1998). e skin conductance response is a measure of autonomic arousal and, hence, a body-based measure of
emotion processing (see figure opposite for further details). Beara et al. (1995) report that humans with amygdala damage fail to show this conditioned response, but nevertheless are able to verbally learn the association (“when I saw the blue square I got a sho”), whereas amnesic patients with hippocampal damage show a normal conditioned response, but cannot recall the association. is suggests that the association is stored in more than one place: in the amygdala (giving rise to the conditioned fear response) plus in the hippocampus (giving rise to declarative memories of the association). fMRI studies also show that the amygdala may be important for fearrelated conditioning in social seings in whi participants learn fear associations by wating someone else receive a sho (Olsson & Phelps, 2004).
Amygdala lesions in humans can selectively impair the ability to perceive fear in others but not necessarily the other Ekman categories of emotion (Adolphs et al., 1994; Calder et al., 1996). For example, patient DR suffered bilateral amygdala damage and subsequently displayed a particular difficulty with recognizing fear (Calder et al., 1996). She was also impaired to a lesser degree in recognizing facial anger and disgust. She could imagine the facial features of famous people, but not of emotional expressions. She could recognize famous faces and mat different views of unfamiliar people, but could not mat pictures of the same person when the expression differed (Young et al., 1996). DR also shows comparable deficits in recognizing vocal emotional expressions, suggesting that the deficit is related to emotion processing rather than modality-specific perceptual processes (Sco et al., 1997). While it has been suggested that selective impairments in fear may arise because of a failure to aend closely to the eyes (Adolphs et al., 2005), this cannot account for the fact that some patients fail to recognize fear in spee (Sco et al., 1997) or music (Gosselin et al., 2007).
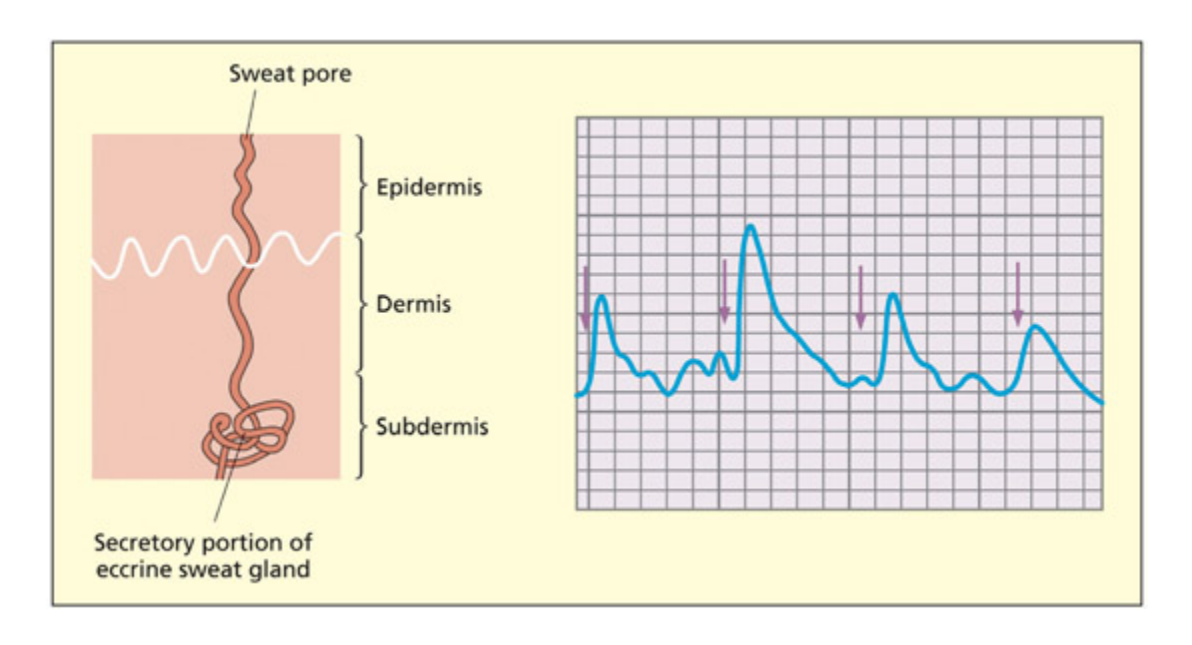
📊 그림 설명
피부 전도 반응(SCR) 측정 방법을 설명하는 그림이다. 손에 부착된 전극을 통해 피부의 전기 전도도 변화를 기록하며, 자율 신경계 흥분에 의한 미세한 발한이 전도도를 증가시킨다. 자극 제시 후 1~5초 사이에 SCR 피크가 나타나며, 이는 감정적 자극에 대한 자율 신경계 반응의 지표로 활용된다.
e skin conductance response (SCR) method involves recording anges in electrical conductivity on a person’s skin on the hand. Heightened arousal can lead to more sweat even without overt sweating taking place. A person’s SCR can be ploed as a continuous trace throughout the experiment. A peak SCR occurs between 1 and 5 s aer face presentation.
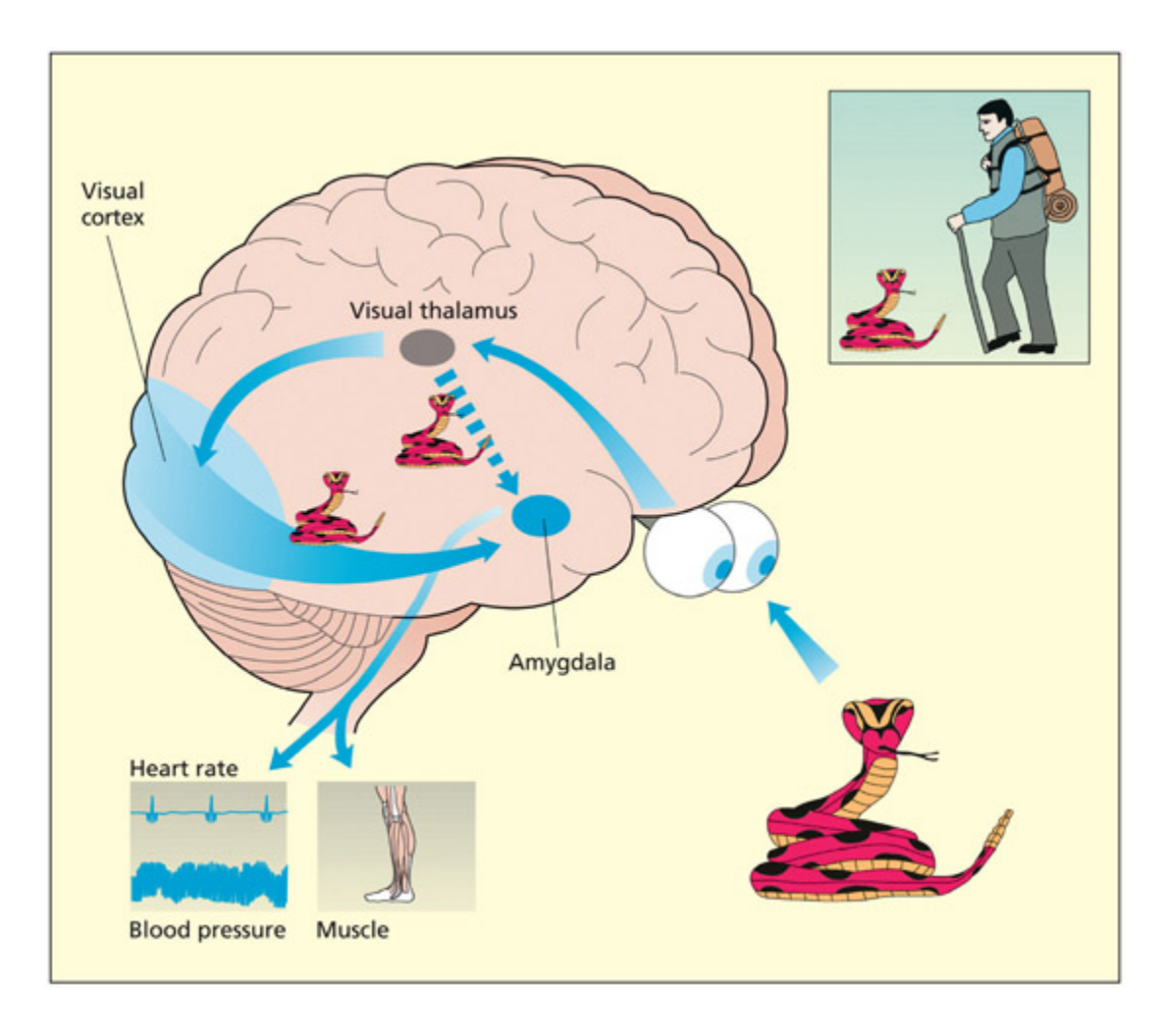
📊 그림 설명
LeDoux가 제안한 편도체의 위협 자극에 대한 빠른 반응 경로를 보여주는 그림이다. 시상에서 편도체로 향하는 빠른 하위피질 경로(low road)와 시각 피질을 경유하는 느린 경로(high road)가 도식화되어 있다. 이 이중 경로 모델은 뱀이나 거미 같은 위협 자극에 대해 의식적 인식 이전에 편도체가 반응할 수 있음을 설명한다.
Le Doux has argued that the amygdala has a fast response to the presence of threatening stimuli su as snakes.
Functional imaging studies generally support, and extend, these conclusions. Morris et al. (1996) presented participants with morphed faces on a happy– neutral–fearful continuum. Participants were required to make male–female classifications (i.e. the processing of emotion was incidental). Le amygdala activation was found only in the fear condition; the happy condition activated a different neural circuit. Winston et al. (2003) report that amygdala activation was independent of whether participants engaged in incidental viewing or explicit emotion judgments. However, other regions, including the ventromedial frontal lobes, were activated only when making explicit judgments about the emotion. is was interpreted as reinstatement of the “feeling” of the emotion.
Some researers have argued that the ability to detect threat is so important, evolutionarily, that it may occur rapidly and without conscious awareness (Le Doux, 1996). Ohman et al. (2001) report that people are faster at detecting snakes and spiders among flowers and mushrooms than the other way around, and that sear times are suggestive of preaentive “popout” (see Chapter 7). When spiders or snakes are presented subliminally to people with spider or snake phobias, then participants do not report seeing the stimulus but show a skin conductance response indicative of emotional processing (Ohman & Soares, 1994). In these experiments, aranophobic participants show the response to spiders, not snakes; and ophidiophobic participants show a response to snakes, but not spiders. In terms of neural pathways, it is generally believed that there is a fast subcortical route from the thalamus to the amygdala and a slow route to the amygdala via the primary visual cortex (Adolphs, 2002; Morris et al., 1999). Functional imaging studies suggest that the amygdala is indeed activated by unconscious fearful expressions in both healthy participants (Morris et al., 1999) and in a “blindsight” patient with damage to primary visual cortex (Tamieo et al., 2012). is is consistent with a subcortical route to the amygdala, although it is to be noted that the temporal resolution of fMRI does not enable any conclusions to be drawn about whether the route is fast or slow.
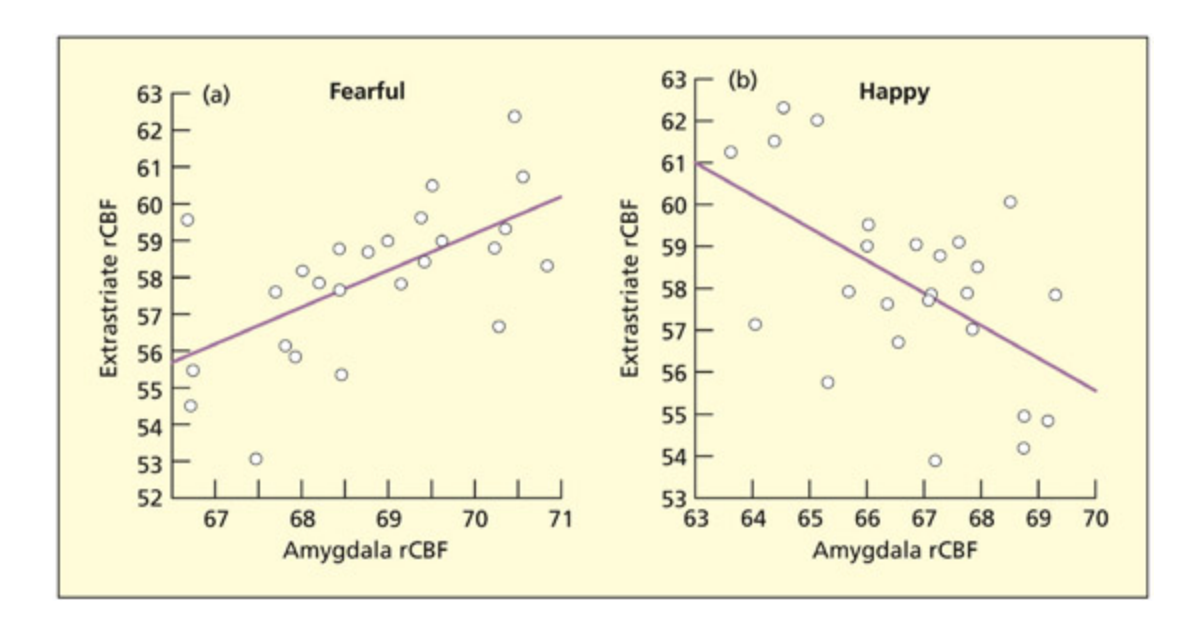
📊 그림 설명
공포 표정과 행복 표정에 대한 편도체와 시각 피질의 혈류 변화를 보여주는 fMRI/PET 결과이다. 공포 표정을 볼 때 편도체의 혈류 증가가 외선조 시각 영역의 활성화와 양의 상관관계를 보이며, 이는 위협 감지 시 시각적 경계를 강화하는 기능을 반영한다. 행복 표정에서는 반대 패턴이 관찰된다.
When shown fearful faces, increases in regional cerebral blood flow in the amygdala (rCBF) are associated with increases in blood flow in extrastriate visual regions involved in recognition of the potential threat. e reverse is true of happy faces.
From Morris et al., 1998. Reprinted by permission of Oxford University Press.
While there is convincing evidence for the role of the amygdala in fear processing it should not be concluded that the amygdala is the “fear center” of the brain. First, fear may depend on a wider network (of whi the amygdala is a key hub). Indeed, it affects the autonomic system (via the hypothalamus) to generate a fight or flight reaction (Le Doux et al., 1988) and it increases activity in visual cortex to promote vigilance to threat (Morris et al., 1998), among others. Second, the fact that the amygdala is a key part of the fear network doesn’t preclude it from being involved in other forms of emotional processing. With regards to learning of stimulus-emotion associations there is evidence that the amygdala is involved in learning positive associ ations, based on food rewards, as well as fear-conditioning (Baxter & Murray, 2002). However, the amygdala system for positive associations operates somewhat differently to fear-conditioning and depends on different nuclei. For example, selective lesions of the amygdala in animals does not affect learning of classically conditioned light-food associations, in whi the animal learns to approa the food cup when the light comes on (Hatfield et al., 1996), although su lesions are known to affect learning that a light predicts a sho. However, amygdala lesions do affect other aspects of reward-based learning su as second order condi tioning in whi a light + tone is subsequently paired with absence of food (aer learning that a light alone predicts food), or learn ing that the food is devalued (Hatfield et al., 1996). Recent functional imaging studies that compare stimuli with learned positive and negative associations relative to emotionally neutral ones but do not rely on facial expressions have revealed amygdala activation to negative and positive affective stimuli; for instance, comparing positive, negative, and neutral tastes (Small et al., 2003), smells (Winston et al., 2005), pictures, and sounds (Anders et al., 2008). However, most fMRI studies do
not have the spatial resolution to reliably distinguish between subregions within the amygdala.
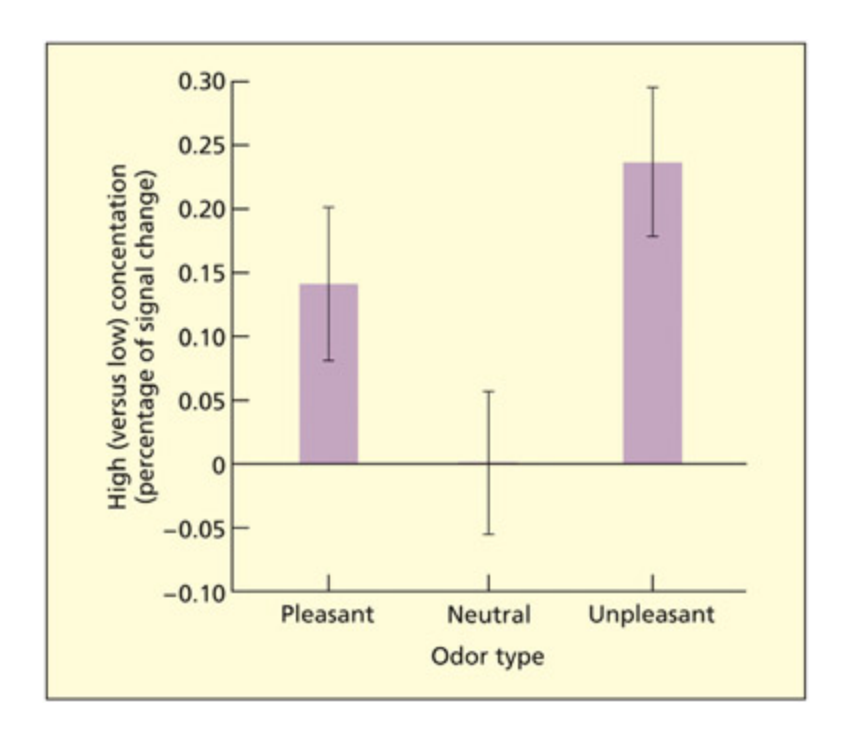
📊 그림 설명
편도체가 쾌적한 냄새와 불쾌한 냄새 모두에 반응하지만, 중립적 냄새에는 반응하지 않음을 보여주는 뇌 영상 데이터이다. 이 결과는 편도체가 단순히 공포에만 특화된 것이 아니라, 정서적 가치를 지닌 자극 전반에 대해 폭넓은 역할을 수행한다는 증거를 제공한다.
ere is evidence that the amygdala responds to pleasant and unpleasant smells (but not neutral smells). is suggests a wider role of the amygdala in emotion processing, in contrast to the commonly held assumption that it is specific to fear.
From Dolan, R. J., 2007. e human amygdala and orbitofrontal cortex in behavioural regulation. Philosophical Transactions of the Royal Society of London Series B , 362, 787–799. Reproduced with permission.
주의
편도체(amygdala)를 단순히 “공포의 중심(fear center)“으로 기억하면 시험에서 감점당할 수 있다. 편도체는 **공포뿐 아니라 긍정적 연합 학습(보상 기반)**에도 관여하며, 쾌적한/불쾌한 냄새, 맛, 소리 모두에 반응한다. 핵심은 편도체가 정서적 가치를 지닌 자극 전반의 학습과 기억에 관여하는 허브라는 것이다. 다만 공포 조건화와 보상 학습은 편도체 내 **서로 다른 핵(nuclei)**에 의존한다.
The insula: disgust and interoception
Summary
인서라(insula)는 측두엽 아래에 위치하며 통증/맛 인식 등 신체 감각 처리에 관여한다. 역겨움(disgust)은 진화적으로 오염/질병 회피와 관련되며, 인서라의 내부 감각(interoception) 기능과 밀접하게 연결된다.
e insula is a small region of cortex buried beneath the temporal lobes (it literally means “island”). It is involved in various aspects of bodily perception including important roles in pain perception and taste perception. e word disgust literally means “bad taste,” and this category of emotion may be evolutionarily related to contamination and disease through ingestion.
Key Terms
Insula
A region of cortex buried beneath the temporal lobes; involved in body perception and contains the primary gustatory cortex; responds to disgust.
인서라 손상(특히 헌팅턴병)은 역겨움 표정 인식에 선택적 장애를 유발하며, fMRI에서 역겨움 표정은 인서라를 활성화하지만 편도체는 활성화하지 않는다. 자신이 역겨움을 느끼거나 타인의 역겨움을 관찰할 때 동일한 인서라 영역이 반응하며, 도덕적 역겨움도 인서라 활성화와 연결된다. 인서라는 감정의 내재적 느낌(feeling)을 형성하는 내부 감각 모니터링 기능을 수행한다.
Patients with Huntington’s disease can show selective impairments in recognizing facial expressions of disgust (Sprengelmeyer et al., 1997) and relative impairments in vocal expressions of disgust (Sprengelmeyer et al., 1996). e degree of the disgust-related impairments in this group correlates with the amount of damage in the insula (Kipps et al., 2007). Selective lesions resulting from brain injury the insula can affect disgust perception more than recognition of other facial expressions (Calder et al., 2000). In healthy participants undergoing fMRI, facial expressions of disgust activate this region, but not the amygdala (Phillips et al., 1997). Feeling disgust oneself and seeing someone else disgusted activates the same region of insula (Wier et al., 2003).
We use the word “disgust” in at least one other context, namely to refer to social behavior that violates moral conventions. Disgusting behavior is said, metaphorically, to “leave a bad taste in the mouth.” But is there more to this than metaphor? Some have argued that moral disgust has evolved out of nonsocial, contamination-related disgust (Tybur et al., 2009). Moral disgust also results in activity in the insula (Moll et al., 2005) and is associated with subtle oral facial expressions aracteristic of disgust more gen erally (Chapman et al., 2009).
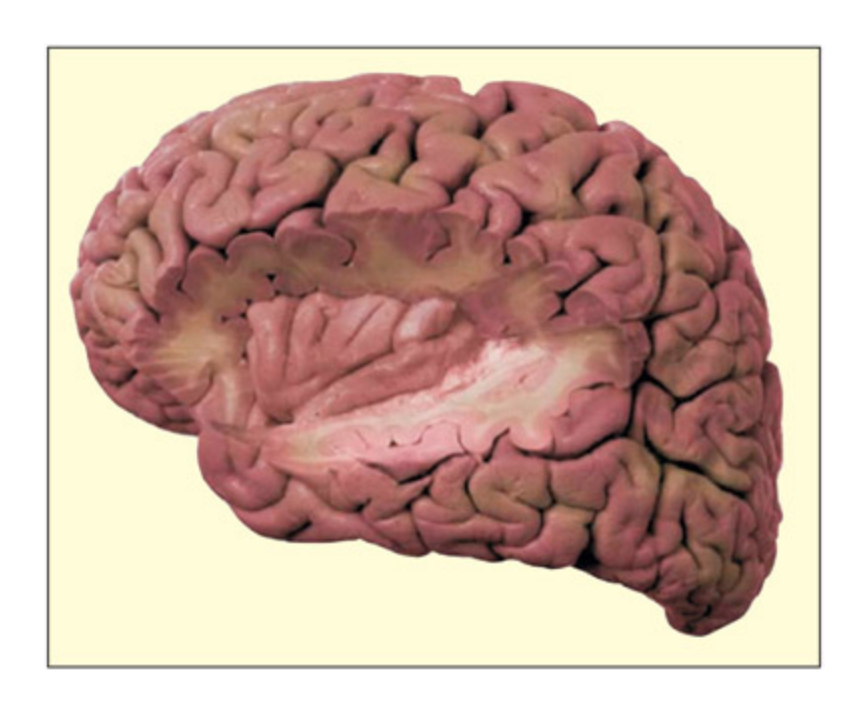
📊 그림 설명
인서라(insula)의 해부학적 위치를 보여주는 뇌 단면도이다. 인서라는 측두엽 아래에 양측으로 위치한 피질 영역으로, 감정과 관련된 신체 감각(내부 감각, interoception)의 형성에 관여한다. 특히 역겨움(disgust) 인식에 핵심적인 역할을 하며, 자신의 역겨움과 타인의 역겨움 관찰 시 동일 영역이 활성화된다.
e insula is an island of cortex lying, bilaterally, underneath the temporal lobes. It is implicated in the creation of bodily feelings associated with emotions, and in the perception of disgust in particular. From Singer et al., 2009. © 2009 Elsevier. Reproduced with permission.
e insula is generally considered to have a wider role in emotional processing, in addition to a more specific involvement in disgust. Specifically, it is regarded as monitoring (probably both consciously and unconsciously) the internal state of the body—a process known as interoception. Bodily reactions are aracteristic of emotions and may constitute the “feeling” of an emotion (Craig, 2009; Singer et al., 2009). is is reminiscent of James-Lange theory, but, whereas that account argued that emotions can be reduced to bodily states (i.e. are synonymous with them), modern accounts assume that they are one aspect of an emotion.
Orbitofrontal cortex: contextualized emotions, and emotional feelings
Summary
안와전두피질(orbitofrontal cortex)은 자극의 현재 가치(보상/처벌)를 맥락에 따라 평가한다. 초콜릿을 반복 섭취하면 활성화가 내측부(pleasant)에서 외측부(unpleasant)로 이동하며, 외측부는 기대와 다른 자극(예: 미소 대신 분노)이나 금전적 손실에도 반응한다.
One general function of the orbitofrontal cortex is in computing the current value of a stimulus, i.e. how rewarding the stimulus is within the current
context. For example, ocolate may be a rewarding stimulus, but it may not be currently rewarding if one is full-up or if eating it may incur the anger of someone else. Small et al. (2001) asked participants to eat ocolate between several blos of functional imaging. Initially, the ocolate was rated as pleasant and participants were motivated to eat it, but the more they ate the less pleasant it became and they were less motivated to eat it. is ange in behavior was linked to anges in activity in orbitofrontal regions. Specifically, there was a shi in activity from medial regions (pleasant/wanting) to lateral regions (unpleasant/not-wanting). Other studies are consistent with different regions of orbitofrontal cortex coding rewards and punishments (e.g. for a review see Kringelba, 2005). For instance, activation of lateral orbitofrontal cortex is found when a rewarding smile is expected, but an angry face is instead presented (Kringelba & Rolls, 2003) and is correlated with amount of monetary loss on a trial (O’Doherty et al., 2001).
Key Terms
Extinction learning
Learning that a previously rewarded stimulus is no longer rewarded.
소멸 학습(extinction learning)은 보상이 중단된 자극에 대해 행동을 조정하는 과정이며, 안와전두피질 손상은 역전 학습과 소멸 학습에 어려움을 유발한다. 안와전두피질의 활성도는 자극의 주관적 즐거움(맛, 음악)과 관련되며, 인지 재평가 시 외측전두피질 활성이 높아지면 안와전두피질과 편도체 활성은 감소한다.
e orbitofrontal cortex may enable flexible anges in behavior to stimuli that are normally rewarding (or recently rewarding) but suddenly cease to be. is can account for its role in reversal learning (in whi rewarded and nonrewarded stimuli are reversed) and extinction (in whi a rewarded stimulus is no longer rewarded). Eating ocolate until it is no longer pleasant can be regarded as a form of extinction. Lesions in these regions in humans lead to difficulties on these tasks, and the amount of
difficulty in reversal learning correlates with the level of socially inappropriate behavior of the patients (Rolls et al., 1994).
Activity in the orbitofrontal cortex has been linked to participants’ subjective reports of pleasantness to stimuli su as tastes (McClure et al., 2004b) and music (Blood & Zatorre, 2001). Importantly, these ratings of pleasantness are not just affected by the stimulus itself, but also the participants’ beliefs about the product. Being told the price of a wine affects ratings of pleasantness upon tasting it—more expensive wines taste nicer and perceived pleasantness was again related to activity in the medial part of the orbitofrontal cortex (Plassmann et al., 2008). Of course, the experimenters administered some of the same wines twice giving the participants different prices so the stimuli were physically identical but their beliefs about the quality of the wine were not identical.

📊 그림 설명
동일한 자극(예: 초콜릿)이 맥락에 따라 쾌감 또는 혐오를 유발할 수 있음을 보여주는 안와전두피질(OFC) 활성화 패턴이다. 초콜릿을 처음 먹을 때는 내측 OFC(쾌감)가 활성화되지만, 포만 상태에서는 외측 OFC(불쾌감)로 활성화가 이동한다. 이는 OFC가 자극의 현재 가치를 맥락적으로 계산하여 유연한 행동을 가능하게 함을 보여준다.
e same stimulus can elicit pleasure or aversion depending on context (e.g. the person’s motivational state). Chocolate is normally pleasant, but if you have just eaten two bars of it you probably do not want any more. e orbitofrontal cortex computes the current emotional status of a stimulus (i.e. whether it is currently desired or not), thus enabling flexible behavior. Other regions in the brain may code the long-term value of a stimulus (i.e. whether it is normally desired or not).
e lateral prefrontal cortex and the orbitofrontal cortex might serve somewhat different functions in regulating and contextualizing emotions. Osner et al. (2002) presented negative images (e.g. of someone in traction in a hospital) to participants in one of two conditions: either passively viewing them or a cognitive condition in whi they were instructed to
reappraise ea image “so that it no longer elicited a negative response.” eir analysis revealed a trade-off between activity in the lateral prefrontal cortex (high when reappraising) and the medial/orbital frontal cortex and amygdala (high during passive looking). When participants are asked to reappraise the stimulus negatively, i.e. making it worse than it looks, then this also engenders a similar network in the lateral prefrontal cortex but tends not to dampen activity in the ventromedial prefrontal cortex and amygdala (Osner et al., 2004).
Anterior cingulate: response evaluation, autonomic responses, and pain
Summary
전대상피질(anterior cingulate cortex)은 행동의 보상/처벌 가치를 평가하며, 안와전두피질이 자극 가치를 계산하는 것과 구분된다. 사이버볼 게임(Cyberball) 실험에서 사회적 배제 조건은 전대상피질 활성화와 주관적 고통 수준의 상관관계를 보였다(Eisenberger et al., 2003).
In the apter on the Executive Brain, it was noted that the anterior cingulate is involved in the detection of errors and monitoring of response conflict su as on the Stroop test. Although this theory is not normally coued in terms of emotional processing (Carter et al., 1998) it can be. Rushworth et al. (2007) argue that the function of the anterior cingulate is to assess the value of responses, i.e. whether an action is likely to elicit a reward or punishment. is may differ from the function of the orbitofrontal cortex whi computes whether a given stimulus is currently rewarded or punished. Male monkeys with anterior cingulate lesions fail to adjust their responses, when reaing for food, when simultaneously shown a dominant male or a female in estrus, whereas most control monkeys will pay close aention to these social stimuli, and hence take longer to respond to the food (Rudebe et al., 2006).
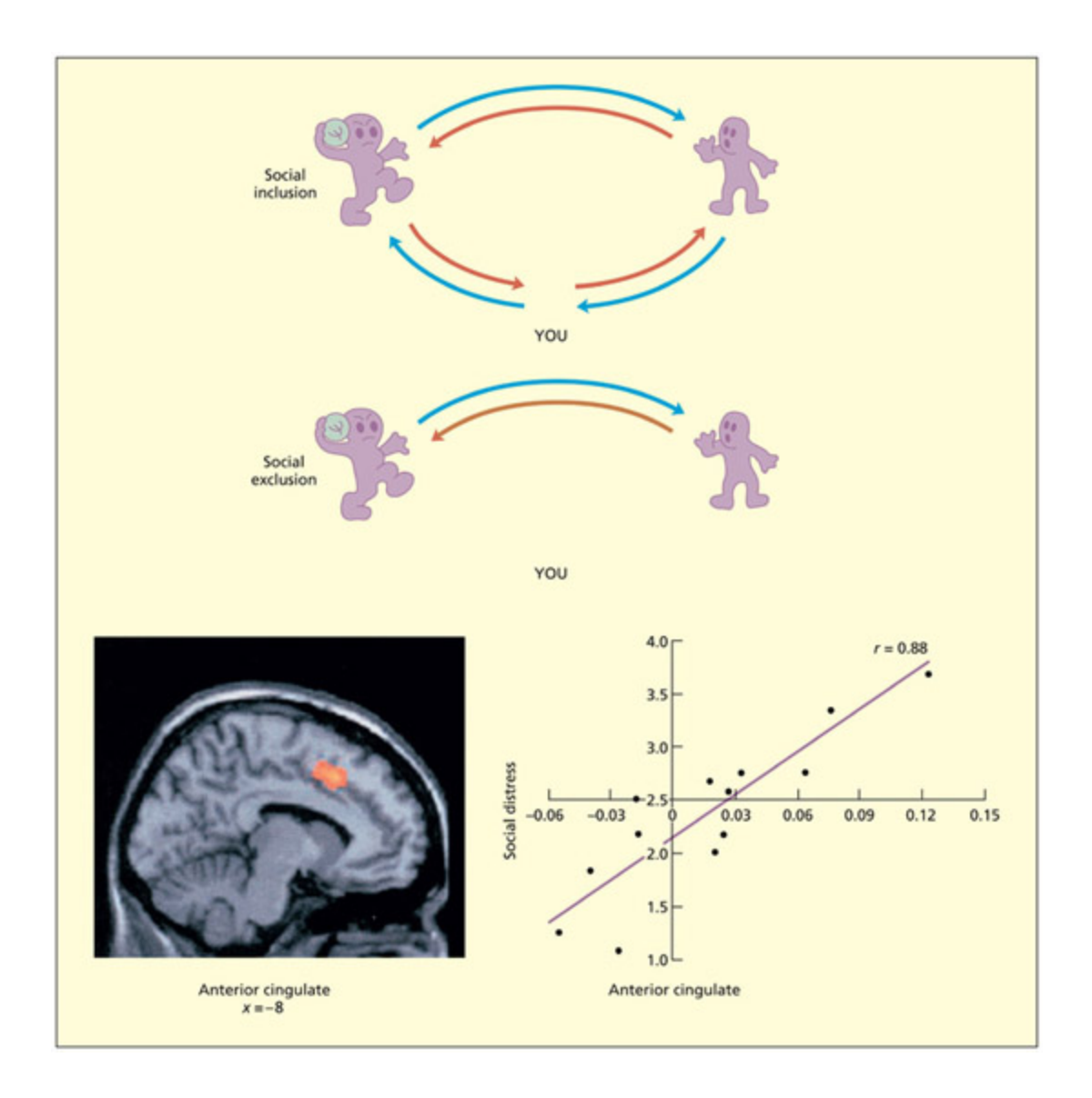
📊 그림 설명
Cyberball 게임 실험의 설계와 결과를 보여주는 그림이다. 참가자가 세 명 중 한 명으로 공 던지기 게임에 참여하며, 사회적 배제 조건에서는 다른 두 참가자가 서로에게만 공을 던진다. fMRI 결과, 사회적 배제 시 전대상피질(anterior cingulate)이 활성화되며, 이 활성화 수준은 주관적 고통 보고와 상관관계를 보인다.
In the Cyberball game a participant must decide whi of two other players to throw the ball to. In a social exclusion condition, two of the players always send the ball to ea other and never to the participant. In a social inclusion condition, all players get to play. Social exclusion tends to activate the anterior cingulate and this correlates with subjective levels of distress. Boom figures from Eisenberger et al., 2003. © 2003 American Association for the Advancement of Science. Reproduced with permission.
From Ward, 2012, p. 195.
Key Terms
Ventral striatum
Part of the basal ganglia that includes the nucleus accumbens; involved in a “limbic circuit” connecting the orbitofrontal cortex, basal ganglia, and thalamus.
복측선조체(ventral striatum)는 기저핵의 일부로, 안와전두피질-기저핵-시상 연결의 변연계 회로 핵심 요소이다. 전대상피질은 인서라와 달리 신체 반응의 출력에 초점을 맞추며, 손상 시 SCR/심박수/혈압 변화가 방해된다. 타인의 통증 관찰 시에도 전대상피질이 활성화되어, 사회적 고통과 신체적 통증이 공통 회로를 공유함을 시사한다.
e anterior cingulate cortex, like the insula, is involved in processing bodily signals that aracterize emotions but, whereas the insula is more concerned with the input (and awareness) of these signals, the anterior cingulate is more concerned with the output of bodily responses. Lesions in this area disrupt the skin conductance response (Tranel & Damasio, 1995) and anges in heart rate and blood pressure (Critley et al., 2003) to emotional stimuli.
e anterior cingulate also receives inputs (via the thalamus) relating to pain, and may regulate feelings of pain via output connections to the periaqueductal gray (a region that is ri in endogenous opioids). As well as responding (e.g. in fMRI) to physically painful stimuli, su as mild electric shos, wating someone else in pain activates some of the same regions (Singer et al., 2004). us it responds to the perception of pain in others as well as to physical pain in oneself. is idea is returned to in later discussions on empathy. It has also been claimed that being separated from a loved one or being socially excluded in general is “painful,” and these more social forms of pain may indeed involve the pain circuitry of the brain. Eisenberger et al. (2003) conducted an fMRI study of a Cyberball game involving three players, including the one person being scanned. Players could opt to throw the ball to one of the two other players. However, aer a while the game was fixed su that two players consistently threw to ea other excluding the person in the scanner. ere were two other conditions: one in whi the player was included, and one in whi they were excluded but given the cover story of “due to tenical difficulties.” Activity in the anterior cingulate correlated with self-reported distress during social exclusion. A region in the prefrontal cortex (right ventro-lateral prefrontal
cortex) was linked to social exclusion, but not exclusion due to “tenical difficulties” whi they interpret as playing a controlling role in limiting the distress of social exclusion.
Ventral striatum and reward
Summary
복측선조체의 측좌핵(nucleus accumbens)은 도파민 신경회로를 통해 보상 예측 오차(reward prediction error)를 부호화하며, 예상보다 높은 보상에는 활성이 증가하고 예상 보상이 빠지면 기저선 이하로 감소한다. 협력을 통해 얻은 보상은 비협력적 상황보다 복측선조체 활성도를 더 높이며, 사회적 고립 보고 개인은 사회적 장면에 대해 낮은 활성을 보인다.
e dorsal region of the striatum has more sensorimotor properties (e.g. involved in habit formation), whereas the ventral region may be more specialized for emotions, although the distinction is relative not absolute (Voorn et al., 2004). ere are several loops that connect regions within the frontal cortex to the basal ganglia and on to the thalamus before returning to the frontal cortex (Alexander & Cruter, 1990). e loops modulate brain activity within these frontal structures and, hence, increase or decrease the probability of a particular behavior. e loop that is of particular relevance to reward-based learning (the “limbic circuit”) starts and ends in the orbitofrontal cortex and limbic regions (including amygdala and anterior cingulate), passing through the basal ganglia (including the ventral striatum) and thalamus.
Neurons containing the neurotransmier, dopamine, project from the midbrain to a region in the ventral striatum called the nucleus accumbens. Psyomotor stimulants su as amphetamine and cocaine may exert their effects via this system (Koob, 1992). Other rewarding stimuli activate this region. Dopamine release in the nucleus accumbens of male rats increases when a female is introduced to the cage, and increases further if they have sex (Pfaus et al., 1990). Neutral stimuli previously associated with food increase the release of dopamine in the nucleus accumbens of rats (Robbins et al., 1989). In humans, an fMRI study shows that the greater the monetary reward that could be obtained in a task the larger the activity in the ventral striatum (Knutson et al., 2001). However, social stimuli are rewarding too, and activity in this region tends to be greater when a reward (e.g. monetary) is obtained via cooperation with another human, than when it is obtained
from noncooperation with a human or cooperation with a nonsocial agent su as a computer (Rilling et al., 2002).
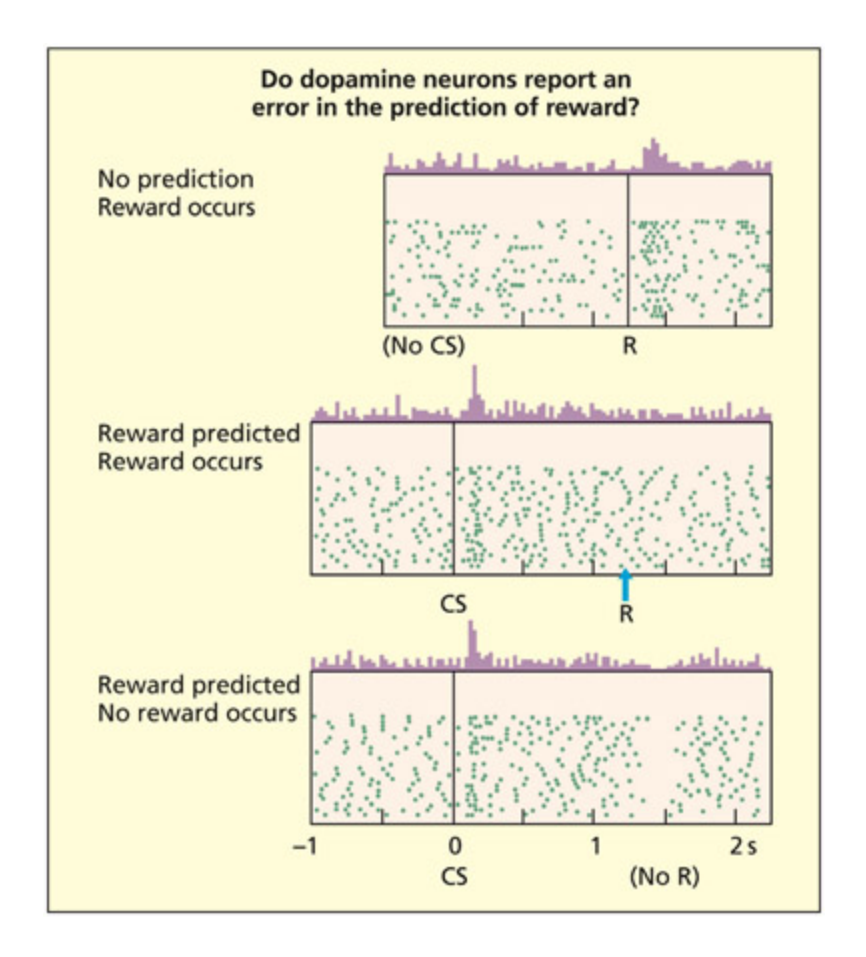
📊 그림 설명
원숭이 복측선조체의 도파민 뉴런 단일세포 기록 결과이다. 예상치 못한 보상(과일 주스)이 주어지면 뉴런이 강하게 반응하고(상단), 보상이 단서로 예측되면 단서에 반응하고 보상에는 반응하지 않으며(중단), 예상된 보상이 생략되면 발화가 기저선 이하로 떨어진다(하단). 이는 도파민 뉴런이 보상 그 자체가 아닌 보상 예측 오차(reward prediction error)를 부호화함을 보여준다.
Single-cell recordings of dopamine neurons in the ventral striatum of monkeys show that the neuron responds when an unexpected reward of fruit juice is given (top), but if the reward is predicted by a cue (the conditioned stimulus) then the neuron responds to the cue and not the reward (middle). If an expected reward is omied (boom) the firing of the neuron falls below baseline. e results suggest that these neurons code the difference between the predicted reward and actual reward, rather than reward itself.
From Sultz et al., 1997. © 1997 American Association for the Advancement of Science. Reproduced with permission.
One contemporary idea is that these dopaminergic neurons are not encoding reward per se, but the difference between the predicted reward and actual reward (e.g. Sultz et al., 1997). Aer training to perform an action when presented with a light or tone cue, dopaminergic neurons in monkeys
eventually respond to the conditioned cue itself rather than the subsequent reward (Sultz et al., 1992). If no subsequent reward appears then their activity drops below baseline, indicating that a reward was expected. Some fMRI studies of decision making in humans also suggest that activity in the ventral striatum is greater when a reward is beer than expected, rather than when a reward is high per se (Hare et al., 2008). Self-reported lonely people show less activity in the ventral striatum when shown photos of social scenes (relative to non-lonely people), arguably because they predict them to be less rewarding (Cacioppo et al., 2009).
Evaluation
is section has outlined a set of regions that are critically involved in the processing of emotions. In social animals, su as humans, these emotional brain regions play a key role in evaluating and judging social stimuli. For instance, the amygdala is not only implicated in evaluating whether a tone will lead to a sho, but also in evaluating whether another person is afraid; the anterior cingulate responds not only to physical pain but also responds to social pain relating to separation and social exclusion; and the nucleus accumbens responds not only to basic rewards (food, sex) but also responds when we opt to cooperate with another person.
e different regions of the emotional brain serve different functions, and this is at odds with earlier theories of emotion (e.g. the Papez circuit and Maclean’s “limbic brain”). However, there is not a simple one-to-one mapping between brain structure and emotional category (e.g. amygdala = fear, insula = disgust) as predicted by a strong version of the “basic emotion” approa. Of course, “basic emotions” could still be said to exist at the level of brain circuits connecting specialized sub-regions and the best documented examples in the literature are fear and disgust. Other contemporary theories postulate the notion of “core affect” whi consists of perceived internal body states and organized around the dimensions of intensity and pleasantness (Feldman Barre, 2006). While there are brain regions that
appear to have these aracteristics (e.g. the amygdala shows some evidence of traing intensity) on closer inspection their functioning is far more complex (e.g. the evidence that the amygdala has somewhat different roles in fear versus reward conditioning).
Key Terms
Conspecific
Other members of the same species.
동종(conspecific)은 동일 종의 다른 개체를 가리킨다. 현대 감정 이론은 감정이 인지(사고, 신념, 평가)와 깊이 통합된 상태임을 강조하며, 감정과 인지가 분리되어 있다는 이전 관점과 대비된다.
Finally, almost all contemporary theories of emotion allow for a role of “cognition” by whi is meant thoughts and beliefs, appraisals, control meanisms and the like. is is supported by the neuroscience evidence but stands in contrast to some earlier views that emotion was somehow distinct from cognition (for a summary see Phelps, 2006). An emotion is best regarded as a complex affective state in whi cognitions are intrinsically embedded rather than standing apart.
Reading Faces
Summary
얼굴은 사회적 객체로서 타인의 감정 상태, 의도(시선 방향), 사회적 범주(인종, 성별), 성격 특성(신뢰성) 등의 정보를 전달한다. 이 섹션에서는 표정 인식과 시선 감지에 초점을 맞춘다.
e visual processing of faces has been considered previously (see Chapter 6). However, a face is far more than a visual object—it is also a social object, denoting a conspecific. A face conveys important information about another person’s feeling states (e.g. their current emotion), their intentions (e.g. eye gaze provides some clues), their membership of social categories (e.g. race, gender), and perhaps even their dispositions (e.g. trustworthiness). is section first considers facial expressions followed by gaze detection.
Evaluating race and personality in a face is covered elsewhere (Kubota et al., 2012; Ward, 2012).
Recognizing facial expressions
Summary
Bruce-Young(1986) 모델은 표정 인식의 독립 경로를, Haxby(2000) 모델은 FFA(정체성)와 STS(표정/시선)의 구분을 제안한다. 그러나 뇌 손상 연구에서 표정 인식 장애는 STS가 아닌 안와전두피질/체감 피질 손상과 관련되어, 표정 인식은 편도체(공포), 인서라(역겨움) 등 감정 확장 시스템에 의존함이 밝혀졌다.
e two models of face processing already considered in some detail in Chapter 6 are the cognitive model of Bruce and Young (1986) and the neuroanatomical model of Haxby et al. (2000). Both models assume that extracting socially relevant information from faces (e.g. knowing they are happy) is largely separable from recognizing facial identity (i.e. knowing who the person is). However, the two models make different assumptions as to how this is done. In Bruce and Young’s (1986) model there is a dedicated route for recognizing emotional expressions. is route is also assumed to be different from the meanism needed for tasks su as lip-reading or gaze detection. By contrast, the model of Haxby et al. (2000) makes a broad division between time-invariant representations of a face (needed for facial identity and linked to the fusiform face area FFA) and time-varying representations of a face. e laer is assumed to be needed both for recognizing expressions and for gaze processing, and is linked to the superior temporal sulcus (STS). Both the fusiform face area and the superior temporal sulcus are assumed to be part of the “core system” of face processing (i.e. relatively specialized for faces in particular), but, for expressions, this would additionally involve the “extended system” dealing with emotions (including the amygdala, insula, and so on).
To what extent does the available evidence support these two models? According to Calder and Young (2005) the evidence does not equivocally support either of these models. Brain-damaged patients who are poor at recognizing facial expressions but who are relatively good at recognizing facial identity do exist. However, the lesion sites of these patients tend to be in regions su as orbital and ventromedial frontal lesions (Heberlein et al., 2008; Hornak et al., 1996) or somatosensory regions (Adolphs et al., 2000), but not the superior temporal sulcus as predicted by the model of Haxby et
al. (2000). ere is convincing evidence that the superior temporal sulcus plays an important role in detecting gaze direction and lip-reading (outlined in the next section), but the evidence that it is crucially involved in expression recognition is laing. Difficulties in recognizing facial expressions appears to depend on the integrity of the extended system (to borrow the terminology of Haxby et al., 2000) that is involved in the general processing of emotion and is not specific to faces. Calder and Young (2005) also argue that this is inconsistent with the assumption of a single route for recognizing expressions, as originally postulated by the Bruce and Young (1986) model. Instead they argue that the recognition of particular emotion expressions is divided up among different brain regions that are specialized for different categories of emotion (e.g. the amygdala for fear, insula for disgust) or for emotional experience in general (e.g. the orbitofrontal cortex).
Key Terms
Simulation theory
The theory that we come to understand others (their emotions, actions, mental states) by vicariously producing their current state in ourselves.
이 섹션에서는 타인의 감정, 행동, 정신 상태를 이해하는 데 있어 자기 자신에서의 유사한 상태 재현(vicarious simulation)을 기반으로 한 시뮬레이션 이론(Simulation theory)을 제시한다. 이 이론은 특히 감정 인식에 있어 감각운동 시뮬레이션(sensorimotor simulation) 메커니즘의 역할을 강조하며, 예를 들어 타인의 미소를 보는 순간 자신의 행복 관련 감각경로가 활성화되고, 실제로 미소를 짓기 위한 운동 프로그램도 자동으로 실행될 수 있다고 설명한다. **Oberman et al. (2007)**의 연구에 따르면, 펜을 이빨로 물고 있는 상태는 미소와 동일한 근육을 사용하므로, 이 과제를 수행할 경우 행복 감정의 표정 인식이 방해받는다는 실험 결과가 제시되었다. 또한 전기근육도(EMG) 연구에서는 타인의 표정을 잠시 관찰할 뿐이라도, 자신의 얼굴 근육에 미세한 반응이 발생한다는 점이 밝혀졌으며, 이는 감정 인식에 대한 시뮬레이션 메커니즘의 생리적 근거로 해석된다. 뇌 손상(lesion studies)과 전정 감각 피질(somatosensory cortex)에 대한 반사 자극(TMS) 실험 결과 역시, 감정 표정 인식에 있어 시뮬레이션 메커니즘의 직접적 기여를 지지한다. 이는 감정 처리의 신경과학적 기초와 연결되어, 표정 인식이 단순한 시각적 특성 분석을 넘어 감각운동 경로의 활성화를 통해 이루어질 수 있음을 시사한다.
e theory that we come to understand others (their emotions, actions, mental states) by vicariously producing their current state in ourselves.
Although not specifically discussed by Calder and Young (2005) or Haxby et al. (2000), there is one candidate meanism that could serve as a general system for recognizing expressions, but not identity—namely in terms of sensorimotor simulation (Heberlein & Adolphs, 2007). Simulation theory consists of a collection of somewhat different theories based around a unifying idea—namely that we come to understand others (their emotions, actions, mental states) by vicariously producing their current state on ourselves. With regard to emotions, the claim is that when we see someone
smiling then we also activate our own affective pathways for happiness. Moreover, we may activate the motor programs needed to make us smile (this may make us smile ba, or it may prepare a smile response) and we may simulate what this might feel like in terms of its sensory consequences (e.g. muscle stret and tactile sensations on the face). As su, one could possibly recognize emotions su as happiness, fear, and disgust not just in terms of their visual appearance but in terms of the way that the activate the sensorimotor programs of the perceiver.
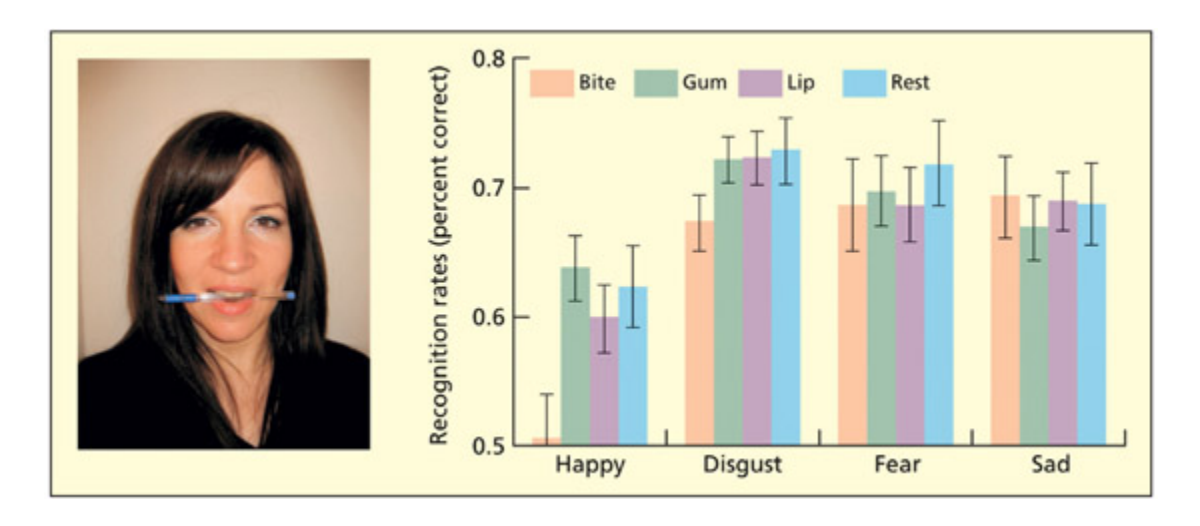
📊 그림 설명
펜을 이빨로 수평으로 물고 있는 과제의 실험 설계와 결과를 보여주는 그림이다. 이 자세는 미소와 동일한 안면 근육을 사용하므로, 미소 관련 감각운동 시뮬레이션을 방해한다. 실험 결과, 이 과제 수행 중 행복 표정의 인식이 선택적으로 저하되어, 표정 인식에 감각운동 시뮬레이션이 기여함을 지지한다.
Placing a pen in the mouth horizontally and holding it with the teeth uses many of the same muscles as smiling. Performing this task can also disrupt recognition of facial expressions of happiness. Data from Oberman et al., 2007.
ere is evidence from electromyographic (EMG) studies that viewing a facial expression produces corresponding tiny anges in our own facial musculature, even if the face is viewed briefly so as to be unconsciously perceived (Dimberg et al., 2000). However, this does not necessarily imply that this is used to recognize expressions. To address this, Oberman et al. (2007) report that biting a pen length-ways uses many of the same muscles involved in smiling. ey subsequently showed that the bite task selectively disrupts the recognition of happiness. Lesion studies (Adolphs et al., 2000) and TMS over the somato sensory cortex (Piter et al., 2008) also suggest a
direct contribution of simulation meanisms to recognizing emotional expressions.
Key Terms
Social referencing
The emotional response of another person may lead to avoidance or interaction with a previously neutral stimulus.
사회적 참조(social referencing)는 타인의 감정 반응이 중립적 자극에 대해 피하기 또는 상호작용을 유도하는 심리적 메커니즘으로, 정서적 적응과 사회적 학습의 핵심 요소이다.
Capgras syndrome
People report that their acquaintances (spouse, family, friends and so on) have been replaced by “body doubles.”
Capgras 증후군은 친족이나 친구 등이 “몸의 이중인물”(body doubles)로 대체되었다고 느끼는 환상적 인식 장애로, 표정 인식과 사회적 참조의 이상과 관련이 있다.
Facial expressions are helpful not only for enabling us to understand what someone else is feeling (e.g. via simulation) but may also be used to modify our own behavior. If human infants are given a novel object, their behavior will be influenced by the response of their primary caregiver—a phenomenon termed social referencing (Klinnert et al., 1983). If the caregiver displays disgust or fear, then the object will be avoided, but if the caregiver smiles, then the ild will interact with the object. is is analogous to the classical conditioning scenario illustrated on p. 383 in whi facial expressions trigger a conditioned response (e.g. fear, happiness) whi becomes associated with the novel object.
”You Look Like My Wife, But You are an Imposter!”
Summary
이 섹션에서는 Capgras 증후군(Capgras delusion) 환자의 피부 전도 반응(Skin Conductance Response, SCR) 특성을 분석하며, 이들이 친숙한 사람의 얼굴을 인식하지만 정서적 반응(emotional response)이 결여되어 “가짜”로 인식하는 현상을 설명한다. 일반적으로 친숙한 얼굴은 SCR을 유발하는 정서적 성분을 포함하지만, Capgras 환자는 이 반응을 보이지 않으며, 이는 Ellis와 Young(1990)의 이론에 의해 설명된다. 이 이론에 따르면 환자는 인식 능력은 유지하지만, 정서적 인식이 결여되어 실제 인물이 아닌 “이중인”(body double)로 판단하게 되며, 이는 특히 가까운 관계자(예: 배우자)에게서 나타나는 현상으로, 이들이 유발하는 정서적 반응의 강도가 가장 크기 때문이다. Tranel 등(1995)의 연구는 일반인에서 친숙한 얼굴에 대한 SCR이 관찰되며, Ellis 등(1997)의 후속 연구는 Capgras 증후군 환자에서 이 반응이 장애(disrupted)됨을 확인했다. 이는 감정 처리와 인식 간의 신경생물학적 연관성을 시사한다.
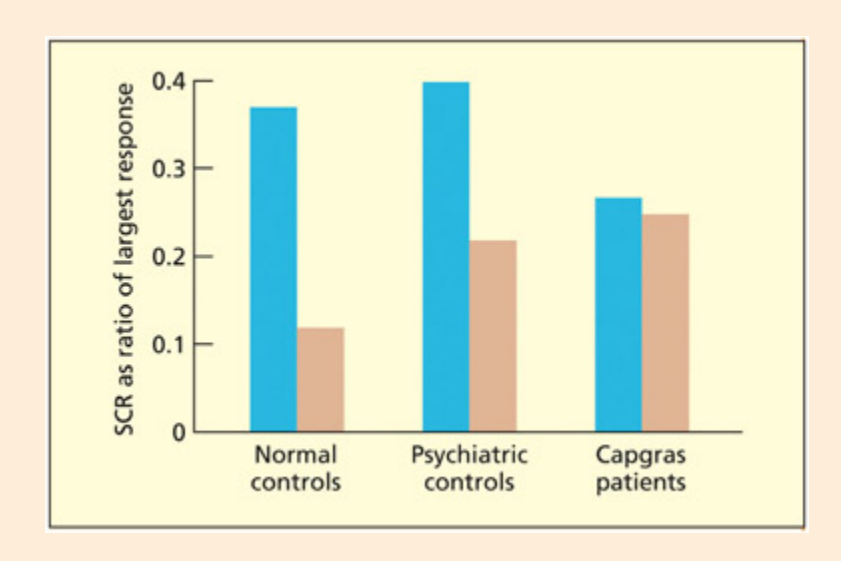
📊 그림 설명
Capgras 증후군 환자와 일반인의 친숙한 얼굴에 대한 피부 전도 반응(SCR) 비교 그래프이다. 일반인은 개인적으로 친숙한 얼굴에 대해 더 큰 SCR을 보이지만, Capgras 환자는 이러한 정서적 반응이 나타나지 않는다. 이는 환자가 얼굴을 시각적으로 인식하면서도 정서적 친밀감을 느끼지 못해 “가짜”로 판단하게 되는 메커니즘을 뒷받침한다.
Most people produce a greater skin conductance response (SCR) to personally familiar faces, but patients with Capgras delusion do not.
Reprinted from Ellis and Lewis, 2001. © 2001 with permission from Elsevier.
In the Capgras syndrome, people report that their acquaintances (spouse, family, friends, and so on) have been replaced by “body doubles” (Capgras & Reboul-Laaux, 1923; Ellis & Lewis, 2001). ey will anowledge that their husband/wife looks like their husband/wife. Indeed, they are able to pi out their husband/wife from a line-up while maintaining all along that he/she is an imposter. To account for this, Ellis and Young (1990) suggest that they can consciously recognize the person, but they la an emotional response to them. As su, the person is interpreted as an imposter. is explains why the people who re doubled are those closest to the patient, as these would be expected to produce the largest emotional reaction. is theory “makes the clear prediction that Capgras patients will not show the normally appropriate skin conductance responses to familiar faces” (Ellis & Young, 1990, p. 244). One general finding in the
neurotypical population is that familiar faces, relative to unfamiliar faces, have an emotional component that reveals itself as a skin conductance response (Tranel et al., 1995). Subsequent resear has confirmed that this skin conductance response response to familiar people is disrupted in Capgras syndrome (Ellis et al., 1997).
Detecting and utilizing eye gaze information
Summary
이 섹션에서는 눈의 시선 정보(eye gaze)가 감정 인식, 사회적 상호작용 및 정서적 적응(emotional adaptation)에 핵심적인 역할을 한다는 점을 강조한다. 특히, 직접적인 시선(direct eye contact)은 영장류에서 감정적 행동(예: 안정화 행동)을 유발하거나 지배적 갈등을 시작하는 중요한 신호로 작동하며, 시선 방향(gaze direction)은 환경에서 중요한 물체에 주의를 집중시키는 기능을 수행한다. 인간의 인지(cognition)에는 타인의 시선 방향을 감지하는 천생의 능력(innate ability)이 존재하며, 이는 신생아부터 관찰되며 학습이 아닌 본능적 반응으로, 상위 측두 세개(superior temporal sulcus)의 신경 활성화와 관련이 있다. 이 부위는 시선 방향 판단 시 활성화되지만, 얼굴 정체성 인식은 융기 얼굴 영역(fusiform face area)에서 처리된다. 자폐증(autism) 환자는 타인의 시선이 자신에게 있는지 감지할 수 있지만, 시선 정보를 통해 행동 예측이나 욕구 추론이 어려운 것으로 나타났다. 예를 들어, ‘차일리가 어떤 초콜릿을 선택할까?‘라는 질문에 답하는 데 실패하며, 이는 공동 주의(joint attention) 부족으로 이어진다. 이러한 차이는 시뮬레이션 이론(simulation theory)과 연결되어, 타인의 정서적 상태를 이해하는 데 있어 시선 정보의 활용이 사회적 능력 발달에 필수적임을 시사한다.
e eye region distinguishes between many emotions, su as smiling or frowning. Moreover, making eye contact can be important for establishing one-to-one communication (dyadic communication), and the direction of gaze can be important for orienting aention to critical objects in the environment. Direct eye contact, in many primates, can be sufficient to initiate emotional behaviors. Macaques are more likely to show appeasement behaviors when shown a direct gaze relative to indirect or averted gazes (Perre & Mistlin, 1990), and dominance struggles are oen initiated with a mutual gaze and terminated when one animal averts its gaze (Chance, 1967).
Baron-Cohen argues that an “eye direction detector” is an innate and distinct component of human cognition (Baron-Cohen, 1995a; Baron-Cohen & Cross, 1992). Babies are able to detect eye contact from birth, suggesting that it is not a learned response (Farroni et al., 2002). is ability is likely to be important for the development of social competence, because the eyes code relational properties between objects and agents (e.g. “mummy sees daddy,” “mummy sees the box”). e superior temporal sulcus contains many cells that respond to eye direction (Perre et al., 1985), and lesions in this area can impair the ability to detect gaze direction (Campbell et al., 1990). Functional imaging studies show that when participants are asked to make judgments about eye gaze (deciding whether the face is looking in the same direction as the last face) then activity is increased in the superior temporal sulcus, but not the fusiform face area (Hoffman & Haxby, 2000). In
contrast, when participants are asked to make judgments about face identity (deciding whether the face is the same as the last one presented) then activity is increased in the fusiform face area, but not the superior temporal sulcus.

📊 그림 설명
자폐 아동의 시선 정보 활용 능력을 테스트한 실험 자료이다. 상단에서 자폐 아동은 누가 자신을 바라보는지 감지할 수 있지만, 하단의 과제에서 Charlie의 시선 방향으로부터 “어떤 초콜릿을 선택할 것인가?”를 추론하는 데 실패한다. 이는 시선 감지 능력은 보존되지만, 시선으로부터 욕구나 의도를 추론하는 능력이 손상됨을 보여준다.
Children with autism are able to detect whi person is looking at them (top), but are unable to infer behavior or desires from eye direction (boom). For example, they are impaired when asked “whi ocolate will Charlie take?” or “whi one does Charlie want?.”
Top photo from Baron-Cohen and Cross, 1992. Reprinted with permission of Blawell Publishing. Boom panel from Baron-Cohen et al., 1995. Reproduced with permission from British Journal of Developmental Psychology. © British Psyological Society.
Children with autism can detect whether the eyes of another person are directed at them and, as su, do not appear to be impaired in the perception of gaze (Baron-Cohen et al., 1995). ey do, however, have difficulties in using gaze information to predict behavior or infer desire. In the four sweets task, a cartoon face of Charlie directs his gaze to one of the sweets. Children with autism are unable to decide: “whi ocolate will Charlie take?” or “whi one does Charlie want?.” e difficulty in utilizing gaze information manifests itself as an absence of joint aention in the social interactions of autistic people (Sigman et al., 1986).
Evaluation
Key Terms
Theory-of-mind
The ability to represent the mental states of others (e.g. their beliefs, desires, intentions).
Theory-of-mind는 타인의 의도, 바람, 믿음 등의 정신 상태를 인식하고 표현하는 능력으로, 멘탈라이징 과정의 핵심 요소이다.
Empathy
The ability to appreciate others’ points of view and share their experiences.
공감(Empathy)은 타인의 관점을 이해하고 경험을 공유하는 능력으로, 감정 처리 관련 뇌 영역과 감각운동 시뮬레이션을 통해 구현된다.
Recognizing facial expressions may depend on several meanisms in the brain. Expressions may be recognized using regions of the brain specialized for emotional processing (including the amygdala and orbitofrontal cortex).
However, sensorimotor simulation may also contribute to recognizing expressions. e superior temporal sulcus is important for recognizing eye gaze and facial/bodily movements, but it is presently unclear whether it is critically involved in expression recognition. e recognition of eye gaze provides important clues about the intentions of others and may interface with other regions involved in making mentalizing inferences.
Reading Minds
Summary
이 섹션에서는 얼굴 표정이 타인의 정신 상태(mental states)인 지식, 신념, 감정, 의도, 욕구 등의 외부적 표현일 뿐이며, 이를 통해 타인의 행동을 예측하는 능력이 진화적으로 발달했다고 설명한다. 시뮬레이션 이론(simulation theory)은 타인의 정신 상태를 이해하기 위해 자신의 감각-운동 경로를 통해 상태의 미러링(mirroring)을 통해 유추하는 방식을 제시하며, 이는 미러 뉴런(mirror neurons)과 연관된 신경 메커니즘을 기반으로 한다. 반면, 이론적 사고(theory-of-mind)는 타인의 정신 상태를 추론하고 추론하는 인지적 과정으로, 자신의 정신 상태와 다른 타인의 상태를 구분할 수 있는 능력을 포함한다. 예를 들어, “당신은 상자에 있다고 생각하지만, 저는 바구니에 있다고 안다”와 같은 상황을 설명할 수 있으며, 이는 일부 이론에서는 도메인 특화 모듈(domain-specific module)을 통해 처리한다고 보기도 한다. 멘탈라이징(mentalizing)이라는 용어는 이론적 사고와 동일한 개념을 설명하지만, 특별한 메커니즘을 암시하지 않는다는 점에서 차별화된다. 이 섹션에서는 미러링(mirroring)과 멘탈라이징(mentalizing) 간의 논쟁이 이 분야의 핵심 이해를 위한 가장 명확한 접근 방식이라고 강조하며, 이는 이전 섹션에서 다룬 감정 처리와 정서적 적응 메커니즘과 연결된 신경과학적 기초를 공유한다.
Facial expressions are only an outward manifestation of someone’s unobservable mental state. e term mental states is used to refer to knowledge, beliefs, feelings, intentions, and desires. Being able to know the content of someone’s mind is a good way of predicting their behavior. Humans, and other species, have evolved meanisms for dealing with this. One meanism that has already been toued upon is simulation. Simulation eories have in common the basic idea that we understand others through a self-centered approa. is may be aieved by a mirroring of states: for instance, seeing you afraid, makes me afraid (by activating my fear-related circuits), and this enables me to infer your mental state. e most common version of simulation theory is linked to perception-action coupling and with the candidate neural meanism being mirror neurons (Gallese, 2001, 2003; Gallese & Goldman, 1998). e other main explanation suggests that there is a meanism for inferring and reasoning about the mental states of others that is commonly referred to as theory-of-mind (Denne, 1978). One hallmark of theory-of-mind is that it enables the representation of different states of mind to one’s own (e.g. “you think it is in the box, I know it is in the basket”)—this is not straightforward for most simulation theories to explain. In some stronger accounts, it is assumed that there is a domain-specific module in the brain for theory-ofmind. e term mentalizing is used by some researers instead of theoryof-mind to denote essentially the same thing, but without carrying the
connotation that it may be a special meanism. Although there are more nuanced theories, the debate between mirroring versus mentalizing offers the clearest way of understanding this literature.
Empathy, mirroring, and simulation theory
Summary
이 섹션에서는 공감(empathy)을 타인의 감정에 대한 감정적 반응 또는 이해로 정의하며, 실험적 맥락에서는 타인의 고통을 유발하는 자극(예: 고통받는 사람의 이미지)을 제시하고, 뇌 활동, 주관적 보고, 신체 반응 등 다양한 방식으로 반응을 측정한다. 또한, 개인 간 공감 능력의 차이(예: 타인의 감정에 얼마나 공감하는가)는 주로 설문지(예: Davis, 1980)를 통해 평가된다. 이론적으로 공감은 미러링(mirroring) 또는 멘탈라이징(mentalizing) 메커니즘과 관련될 수 있지만, 이론적 사고(theory-of-mind) 연구와는 차이가 있다. 이론적 사고 연구는 타인의 정신 상태(예: “Sally는 무엇을 생각하는가?“)를 직접 탐색하는 반면, 공감 연구는 일반적으로 정신 상태에 대한 직접적인 탐색을 피한다.
Empathy refers, in the broadest sense, an emotional reaction to (or understanding of) another person’s feelings. In experimental seings, empathy is oen studied by presenting a stimulus relating to one person (e.g. an image or description of someone in distress) and measuring their response in various ways (brain activity, subjective report, bodily response). It is also possible to measure indi vidual differences in empathy, i.e. the tendency for different people to respond empathically, and this is most frequently done via questionnaire (Davis, 1980). From first principles, empathy could be related to either mirroring or mentalizing meanisms or both. However, resear on empathy typically differs from that done with theory-of-mind in that the laer tends to directly probe knowledge of mental states (e.g. what does Sally think?”), whereas studies on empathy tend not to.
Key Terms
Mirror systems
Neural circuits or regions that disregard the distinction between self and other.
미러 시스템은 자가와 타인의 구분 없이 동작, 감각, 감정을 공유하는 뇌의 신경 회로이다. Iacoboni(2009)는 동작의 미러 시스템이 공감을 지원하기 위해 다른 뇌 영역과 연계될 수 있다고 주장한다.
Iacoboni (2009) has argued that the mirror system for action may be coopted by other regions of the brain to support empathy. Carr et al. (2003) examined a possible link between empathy and action perception/production using fMRI in humans. ey showed participants emotional facial expressions under two condi tions: observation versus deliberate imitation. ey found increased activation for the imitation condition relative to observation in classical mirror-system areas su as the premotor cortex. In addition, they found increased activation in areas involved in emotion su as the amygdala and insula. eir claim was that imitation activates shared motor representations between self and other, but, crucially, there is a second step in whi this information is relayed to limbic areas via the insula. is action-to-emotion route was hypothesized to underpin empathy.
Simulation theories extend the notion of a mirror neuron (see Chapter 8) not only to action, but also to sensation (su as pain and tou) and emotion. e term mirror system is used to convey the idea of neural circuits that disregard the distinction between self and other, but need not necessarily imply action-coding mirror neurons. For example, the insula region is activated both when we are disgusted and when we look at someone else scruning up their face in an expression of disgust (Phillips et al., 1997). Moreover, people who score higher on questionnaire measures of empathy show greater activation of their own disgust regions when wating other people being disgusted (Jabbi et al., 2007). is suggests that we may, in some literal sense, share the emotions of the people around us.

📊 그림 설명
타인의 고통을 관찰할 때 공감적 반응을 유발하는 이미지의 예시이다. 이러한 이미지를 볼 때 관찰자의 뇌에서 물리적 통증 인식에 관여하는 영역(전대상피질, 인서라)이 활성화된다. 이는 시뮬레이션 이론의 핵심 근거로, 타인의 상황을 자신의 경험으로 시뮬레이션하여 공감이 이루어짐을 시사한다.
Do you empathize with someone by simulating how you would feel in their situation? An image su as this one tends to activate parts of the brain involved in the physical perception of pain. © Image Source/Corbis.
Singer and colleagues (2004) investigated empathy for pain. e brain was scanned when anticipating and wating a loved-one suffer a mild electric sho. ere was an overlap between regions activated by expectancy of another person’s pain and experiencing pain oneself, including the anterior cingulate cortex and the insula. In a follow-up to this study, participants in an fMRI scanner wated electric shos delivered to people who were considered either good or bad on the basis of whether they had played fairly or unfairly in a game (Singer et al., 2006). While participants empathically activated their own pain regions when wating the “goodie” receive the electric sho, this response was aenuated when they saw the “baddie” receiving the sho. In fact, male participants oen activated their ventral striatum (linked to beer than expected rewards) when wating the baddie receive the sho—i.e. the exact opposite of simulation theory. is brain activity correlated with their reported desire for revenge. is suggests that, although simulation may tend to operate automatically, it is not protected from our higher order beliefs. Other resear has shown that pain-related regions are activated differently when wating someone in pain depending on whether one takes a self-centered or other-centered perspective and
depending on one’s beliefs about whether the pain was necessary (Lamm et al., 2007). is suggests a significant amount of flexibility in mirroring that some simple versions of simulation theory would not predict.
주의
미러 뉴런(mirror neuron)에 대한 흔한 오해: 미러 뉴런이 공감의 전부를 설명한다는 주장은 과장이다. Singer et al.(2006)의 실험에서 “나쁜 사람”이 전기 충격을 받을 때는 공감 반응이 감소하고, 남성 참가자는 오히려 복수 관련 보상 회로(nucleus accumbens)가 활성화되었다. 이는 미러링이 고차원적 신념과 사회적 판단에 의해 조절됨을 보여주며, 단순한 자동적 시뮬레이션 이론의 한계를 드러낸다.
In order to capture the fact that mirroring does not always occur, some researers have argued that empathy should be understood in terms of the coordinated operation of different kinds of processes. Some researers argue for a division between cognitive empathy and affective empathy for whi the former is effectively synonymous with theory-of-mind and the laer with simulation theory (Baron-Cohen & Wheelwright, 2004; Shamay-Tsoory et al., 2009). Decety and Jason (2004; 2006) offer a different model of empathy comprising of three meanisms. e first relates to simulation but is assumed to reflect a property of many brain systems (rather than converging on action-based mirror neurons). e second relates to a meanism for recognizing self and other as distinct, and this is assumed to be related to the temporoparietal junction area (a key region for theory-ofmind, discussed later). is region responds more when participants are asked to imagine someone else’s feelings and beliefs compared with their own (Ruby & Decety, 2004). A third meanism relates specifically to deliberate efforts to shi perspectives between self and other, and is related to executive functioning (and the lateral prefrontal cortex).
📊 그림 설명
공정/불공정한 상대방이 전기 충격을 받을 때의 뇌 활성화 성별 차이를 보여주는 fMRI 결과이다. 여성(핑크)과 남성(파랑) 모두 불공정한 상대의 충격 시 인서라의 통증 관련 활성이 감소하지만, 남성은 추가로 측좌핵(nucleus accumbens) 활성이 증가하며 이는 복수 욕구와 상관관계를 보인다. 이는 시뮬레이션이 고차원적 신념과 사회적 판단에 의해 조절됨을 보여준다.
Females (pink) and males (blue) show reduced activity in brain regions that respond to pain when wating an unfair player receive a sho (shown here for the insula). In males, activity in the nucleus accumbens, measured while the unfair player received a sho, correlates with their self-reported desire for revenge.
From Singer et al., 2006. © 2006 Nature Publishing Group. Reproduced with permission.
Mind-reading in autism
Summary
이 섹션에서는 자폐 스펙트럼 장애와 관련된 사회적 인지(social cognition)의 이상을 탐구하며, 5세 소년 도널드(Donald)의 사례를 통해 자폐 스펙트럼 장애의 특징적인 행동 패턴을 설명한다. 도널드는 반복적인 손가락 움직임, 물체 회전에 대한 강한 흥미, 타인과의 상호작용 회피, 그리고 외부 자극에 대한 감정적 반응 부족 등의 특징을 보였으며, 이는 사회적 참조(social referencing) 및 이론적 사고(theory of mind)의 결여를 시사한다. 이 사례는 Leo Kanner(1943)가 자폐 스펙트럼 장애의 용어를 처음 제안한 계기였으며, Hans Asperger(1944)의 연구를 통해 아스퍼거 증후군(Asperger’s syndrome)이라는 변형된 형태로 구분되었다. 섹션에서는 자폐 스펙트럼 장애가 정서적 적응(emotional adaptation)과 공감(empathy) 능력에 중대한 영향을 미치며, 이는 앞서 논의된 멘탈라이징(mentalizing) 및 시뮬레이션 이론(simulation theory)의 적용에서 중요한 제한 요소로 작용한다고 분석한다. 특히, 자폐 스펙트럼 장애 환자의 타인의 감정 상태 인식(recognition of others’ emotional states) 및 의도 추론(intentional understanding)의 장애는 사회적 상호작용의 핵심 기능인 미러링(mirroring) 과정의 약화와 관련이 있으며, 이는 정서적 뇌(emotional brain)의 신경회로 이상으로 해석될 수 있다.
He wandered about smiling, making stereotyped movements with his fingers, crossing them about in the air. He shook his head from side to side, whispering or humming the same three-note tune. He spun with great pleasure anything he could seize upon to spin … When taken into a room, he completely disregarded the people and instantly went for objects, preferably those that could be spun … He angrily shoved away the hand that was in his way or the foot that stepped on one of his blocks.
(This description of Donald, aged five, was given by Leo Kanner (1943), who also coined the term “autism.” The disorder was independently noted by Hans Asperger (1944), whose name now denotes a variant of autism).
Key Terms
Autism
The presence of markedly abnormal or impaired development in social interaction and communication, and a markedly restricted repertoire of activities and interests.
자폐 스펙트럼 장애(Autism)는 사회적 상호작용과 의사소통의 심한 이상, 활동 및 흥미 범위의 제한을 특징으로 하며, 미러 시스템 기능 이상 및 이론적 사고(theory of mind) 결손과 연관된다.
Asperger syndrome
Autism with no significant delay in early language and cognitive development.
아스퍼거 증후군은 언어 및 인지 발달에 유의미한 지연이 없는 자폐 스펙트럼 장애의 한 형태이다.
False belief
A belief that differs from one’s own belief and that differs from the true state of the world.
가짜 믿음은 타인의 믿음이 물리적 현실과 다를 수 있다는 인지 능력으로, 이론적 사고(theory of mind)의 핵심 요소이다. 자폐 아동은 Sally-Anne 테스트에서 이 능력의 결손을 보인다.
Autism has been formally defined as “persistent deficits in social communication and social interaction across multiple contexts” and “restricted, repetitive paerns of behavior, interests, or activities” (the American Psyiatric Association, 2013, Diagnostic and Statistical Manual;
DSM-V). It is a severe developmental condition that is evident before 3 years of age and lasts throughout life. ere are a number of difficulties in diagnosing autism. First, it is defined according to behavior because no specific biological markers are known (for a review, see Hill & Frith, 2003). Second, the profile and severity may be modified during the course of development. It can be influenced by external factors (e.g. education, temperament) and may be accompanied by other disorders (e.g. aention deficit and hyperactivity disorder, psyiatric disorders). As su, autism is now viewed as a spectrum of conditions spanning all degrees of severity. It affects 1.2 percent of the ildhood population, and is three times as common in males (Baird et al., 2006). Asperger syndrome falls within this spectrum, and is oen considered a special subgroup. e diagnosis of Asperger syndrome requires that there is no significant delay in early language and cognitive development. e term is more commonly used to denote people with autism who fall within the normal range of intelligence. Learning disability, defined as an IQ lower than 70, is present in around half of all cases of autism (Baird et al., 2006).
Mu of the behavioral data has been obtained from high-functioning individuals in an aempt to isolate a specific core of deficits. One candidate deficit is the ability to represent mental states (Baron-Cohen, 1995b; Fodor, 1992). e first empirical evidence in favor of this hypothesis came with the development of a test of false belief, devised by Wimmer and Perner (1983) and tested on autistic ildren by Baron-Cohen et al. (1985). In the version used with autistic ildren, the Sally–Anne task, the ild is introduced to two aracters, Sally and Anne. Sally puts a marble in a basket so that Anne can see. Anne then leaves the room, and Sally moves the marble to a box. When Anne enters the room, the ild is asked: “Where will Anne look for the marble?” or “Where does Anne think the marble is?“. Children with autism reply: “In the box”; whereas normal ildren (aged 4+) and learningdisabled controls reply: “In the basket.” e erroneous reply is not due to a failure of memory, because the ildren can remember the initial location. It is as if they fail to understand that Anne has a belief that differs from
physical reality—that is, a failure to represent mental states. is has also been called “mind-blindness” (Baron-Cohen, 1995b).
A number of other studies have pointed to selective difficulties in mentalizing compared with carefully controlled conditions. For example, people with autism can understand false photographs, but not false beliefs (Leekam & Perner, 1991); can sequence behavioral pictures, but not mentalistic pictures (Baron-Cohen et al., 1986); are good at sabotage, but not deception (Sodian & Frith, 1992); and tend to use desire and emotion words, but not belief and idea words (Tager-Flusberg, 1992). In all instances, the performance of people with autism is compared with mental-age controls to establish that the effects are related to autism and not general level of functioning.
📊 그림 설명
Sally-Anne 과제의 절차를 도식화한 그림이다. Sally가 구슬을 바구니에 넣고 방을 나간 사이 Anne이 구슬을 상자로 옮기며, “Anne이 돌아오면 어디에서 구슬을 찾을까?”라는 질문이 주어진다. 자폐 아동은 물리적 현실(상자)에 기반하여 답하는 반면, 일반 아동(4세 이상)은 Anne의 거짓 믿음(바구니)을 이해하여 올바르게 답한다.
e Sally–Anne task requires an understanding of “false belief” and tends to be failed by ildren with autism.
Adapted from Wimmer and Perner, 1983.
시험 팁
Sally-Anne 과제를 이해하는 핵심: 자폐 아동이 “상자”라고 답하는 이유는 기억력 문제가 아니다(초기 위치도 기억함). 문제는 타인이 자신과 다른 믿음을 가질 수 있다는 것을 표상하지 못하는 것이다. 이를 “mind-blindness(마음 맹)“라 한다. 단, 이 이론만으로는 자폐의 인지적 강점(예: 숨은 그림 찾기 우수 수행, 체계화 능력)을 설명할 수 없다는 한계가 있다.
e mentalizing or theory-of-mind account of autism has not been without its critics. ese criticisms generally take two forms: either that
other explanations can account for the data without postulating a difficulty in mentalizing (Russell, 1997; Stone & Gerrans, 2006), or that a difficulty with mentalizing is necessary but insufficient to explain all of the available evidence (Frith, 1989).
One difficulty with the theory-of-mind explanation is that it fails to account for cognitive strengths as well as weaknesses. One popular notion of autistic people is that they have unusual gis or “savant” skills, as in the film Rain Man. In reality, these skills are found only in around 10 percent of the autistic population (Hill & Frith, 2003). Nevertheless, some account of them is needed for a full explanation of autism. e unusual skills of some autistic people may be partly an outcome of their limited range of interests. Perhaps one reason why some individuals are good at memorizing dates is that they practice it almost all the time. However, there is also evidence for more basic differences in processing style. For example, people on the autistic spectrum are superior at detecting embedded figures (Shah & Frith, 1983). One explanation for this is in terms of “weak central coherence” (Frith, 1989; Happe, 1999). is is a cognitive style, assumed to be present in autism, in whi processing of parts (or local features) takes precedent over processing of wholes (or global features). A different explanation describes all individuals as having a mix of two different processing styles termed “empathizing” and “systemizing” (Baron-Cohen et al., 2003). Most nonautistic people would lie near the middle and possess a mix of both. People with autism would lie at the extreme systemizing end and la empathizing (Baron-Cohen, 2002). Empathizing allows one to predict another person’s behavior, and care about how others feel. us, a la of empathizing would account for the mentalizing difficulties. Systemizing requires an understanding of lawful, rule-based systems and requires an aention to detail. is would account for their preserved abilities and unusual interests (e.g. in calendar systems).
Key Terms
Broken-mirror theory
An account of autism in which the social difficulties are considered as a consequence of mirror-system dysfunction.
Broken-mirror 이론은 자폐 스펙트럼 장애의 사회적 어려움이 미러 시스템 기능 장애의 결과로 설명된다는 이론이다.
Mu oscillations
EEG oscillations at 8-13 Hz over sensorimotor cortex that are greatest when participants are at rest.
mu 진동(8-13 Hz)은 감각운동 피질에서 휴식 상태에서 가장 두드러지며, 행동 수행/관찰 시 억제(mu suppression)되어 미러 시스템 활동의 지표로 간주된다.
ere are several theories that aempt to explain the social deficits in autism without recourse to a deficit in mentalizing or without postulating the absence of a specialized theory-of-mind meanism. An earlier suggestion is that primary deficit in autism is one of executive functioning (Hughes et al., 1994; Ozonoff et al., 1991; Russell, 1997). For example, on false belief tasks the incorrect answer might be osen because of a failure to suppress the strongly activated “physical reality” alternative. e brokenmirror theory of autism argues that the social difficulties linked to autism are a consequence of mirror-system dysfunction (Iacoboni & Dapreo, 2006; Oberman & Ramaandran, 2007; Ramaandran & Oberman, 2006; Rizzolai & Fabbri-Destro, 2010). Several lines of evidence have been taken to support this. Hadjikhani et al. (2006) found, using structural MRI, that autistic individuals had reduced gray maer in several regions linked to the mirror system, including the inferior frontal gyrus (Broca’s region). Dapreo et al. (2006) conducted an fMRI study in whi autistic ildren and mated controls either observed or imitated emotional expressions. e imitation condition produced less activity in the inferior frontal gyrus of the autistic ildren relative to controls, and this was cor related with symptom severity. Oberman et al. (2005) used EEG to record mu waves over the motor cortex of high-functioning autistic ildren and controls. Mu oscillations occur at
a particular frequency (8–13 Hz) and are greatest when participants are doing nothing. However, when they perform an action there is a decrease in the number of mu waves, a phenomenon termed mu suppression. Importantly, in typical controls mu suppression also occurs when people observe actions and, as su, it has been regarded by some as a measure of mirror-system activity (Pineda, 2005). Oberman et al. (2005) found that the autistic ildren failed to show as mu mu suppression as controls during action observation (wating someone else make a pincer movement) but did so in the control condition of action execution (they themselves make a pincer move ment). Finally, wating someone perform an action increases ones own motor excitability, measured as a motor-evoked potential (MEP) on the body, when TMS is applied to the motor cortex. However, this effect is reduced in autistic people, even though their motor cortex behaves normally in other contexts (eoret et al., 2005).
📊 그림 설명
자폐 스펙트럼 장애에서 관찰되는 숨은 그림 찾기(embedded figures) 과제의 예시이다. 자폐인은 복잡한 도형 속에 숨겨진 단순한 도형을 일반인보다 빠르게 탐지하는 경향을 보인다. 이는 약한 중심 일관성(weak central coherence), 즉 전체보다 부분적 특징 처리를 우선시하는 인지 스타일을 반영한다.
People with autism may be faster at spoing embedded figures similar to the ones shown here.
In sum, there is convincing evidence for mirror-system dysfunction in autism. What is less clear is whether this represents the core deficit and whether it is sufficient to account for the full range of social impairments (including false belief). First, tasks involving imitation and empathy do not rely solely on these kinds of simulation meanisms but also involve deliberate perspective taking, knowledge of social rules, and cognitive control critics (Dinstein et al., 2008; Southgate & Hamilton, 2008). Second, a core deficit elsewhere (e.g. in representing mental states) could nevertheless affect the functioning of the mirror system, and perhaps even lead to structural anges within that system. Heyes (2010) argues that the properties of mirror neurons may be learned as a result of social interactions. So impoverished social interactions may cause mirror-system dysfunction, as well as vice versa.
Is Autism an Extreme Form of the Male Brain?
Summary
이 섹션에서는 자폐 스펙트럼 장애(Autism)가 극단적인 남성형 뇌(extreme male brain)의 형태로 설명될 수 있다는 가설을 제시한다. Baron-Cohen(2002)은 인간의 두뇌 특성을 공감(empathizing)과 시스템화(systemizing) 두 축으로 구분하며, 자폐인은 시스템화 능력(S)이 공감 능력(E)보다 극단적으로 우세한 S >> E 유형을 보인다고 주장한다. 이는 자폐인의 공감 능력 저하(mentalizing difficulties)와 특이한 관심사(unusual interests)를 설명하는 데 기여한다. 남성형 뇌 가설(male brain hypothesis)에 따르면, 자폐의 남성 우위 현상은 남성의 뇌 유형(S > E)이 더 흔하기 때문이며, 유전적 요인도 관련이 있을 수 있다. 특히 터너 증후군(Turner’s syndrome)의 사례에서, 모자에서 유래한 X 염색체(maternal X)를 가진 여성은 자폐 스펙트럼에 더 자주 해당하는 반면, 부모에서 유래한 X 염색체(paternal X)를 가진 여성은 그렇지 않다는 점에서, X 염색체의 유전자(특히 모자 유래)가 자폐에 영향을 줄 수 있음을 시사한다. 이 가설은 아직 인지적 또는 신경학적 메커니즘(cognitive or neural mechanisms)의 실질적 근거를 입증해야 할 과제를 남긴다.
Baron-Cohen (2002) argues that the aracteristics of all individuals can be classified according to two dimensions: “empathizing” and “systemizing.” Empathizing allows one to predict a person’s behavior and to care about how others feel. Systemizing requires an understanding of lawful, rule-based systems and requires an aention to detail. Males tend to have a brain type that is biased toward systemizing (S > E), and females tend to have a brain type that is biased toward empathizing (E > S). However, not all men and women have the “male type” and “female type,” respectively. Autistic people appear to have an extreme male type (S >> E), aracterized by a la of empathizing (this would account for the mentalizing difficulties) and a high degree of systemizing (this would account for their preserved abilities and unusual interests). A questionnaire study suggests that these distinctions hold true (Baron-Cohen et al., 2003). However, it remains to be shown whether these distinctions are merely descriptive
or indeed do reflect two real underlying meanisms at the cognitive or neural level.
One observation that needs to be accounted for is the fact that autism is more common in males. e “male brain hypothesis” offers a simple explanation: males are more likely to have a male-type brain. An alternative, but potentially related, account may be in terms of genetic factors. Whereas it is likely that multiple genes confer autistic susceptibility (Maestrini et al., 2000), one of them may lie on the sexlinked X romosome. Evidence for this comes from Turner’s syndrome. Most men have one X romosome from their mother and a Y romosome from their father (XY). Most women have two X romosomes (XX), one from ea parent. Women with Turner’s syndrome have only a single X romosome (X0) from either their father or mother. If the X romosome has a maternal origin, then the woman oen falls on the autistic spectrum; she does not if it has a paternal origin (Creswell & Skuse, 1999). It is suggested that the paternal X romosome may have a preventive effect on autistic tendencies. An absence of this romosome in some Turner’s women and all normal males (who get their X romosome from their mother) may leave them susceptible to autism.
Neural basis of theory-of-mind
Summary
이 섹션에서는 이론적 사고(theory-of-mind)의 신경 기초를 규명하기 위한 두 가지 주요 증거 출처를 다룬다. 첫째, 일반 참가자에 대한 기능적 영상(functional imaging) 연구와 둘째, 뇌 손상 환자(patients with brain lesions)의 행동 연구이다. 다양한 작업을 통해 정신 상태(mental states)를 추론하는 능력을 평가했는데, 예를 들어 이야기나 만화, 타인과의 상호작용을 통해 정신 상태를 직접 추론하는 작업이 활용되었다. Frith와 Frith(2003)의 리뷰 및 메타분석에 따르면, 멘탈라이징(mentalizing)에 관여하는 3개의 주요 뇌 영역이 확인되었으며, 이는 이론적 사고의 신경 기전을 규명하는 데 핵심적인 역할을 한다. 또한 EEG의 Mu 파동(Mu waves, 8–13 Hz 주파수 범위)은 본인의 행동 수행이나 타인의 행동 관찰 시 감소하는 특성을 보이며, 이는 인간의 미러 뉴런(mirror neurons)에 대한 신경적 지표로 해석될 수 있다. 특히 자폐 스펙트럼 장애(autism spectrum disorder) 아동은 타인의 손동작을 관찰할 때 Mu 파동 억제(mu suppression)가 줄어드는 경향을 보이며, 이는 미러 시스템(mirror system) 기능 장애를 암시하는 Broken-mirror 이론을 지지하는 증거로 제시된다.
Evidence for the neural basis of theory-of-mind has come from two main sources: functional imaging studies of normal participants and behavioral studies of patients with brain lesions. Numerous tasks have been used, including directly inferring mental states from stories (e.g. Fleter et al., 1995), from cartoons (Gallagher et al., 2000) or when interacting with another person (McCabe et al., 2001). A review and meta-analysis of the functional imaging literature was provided by Frith and Frith (2003), who identified three key regions involved in mentalizing.
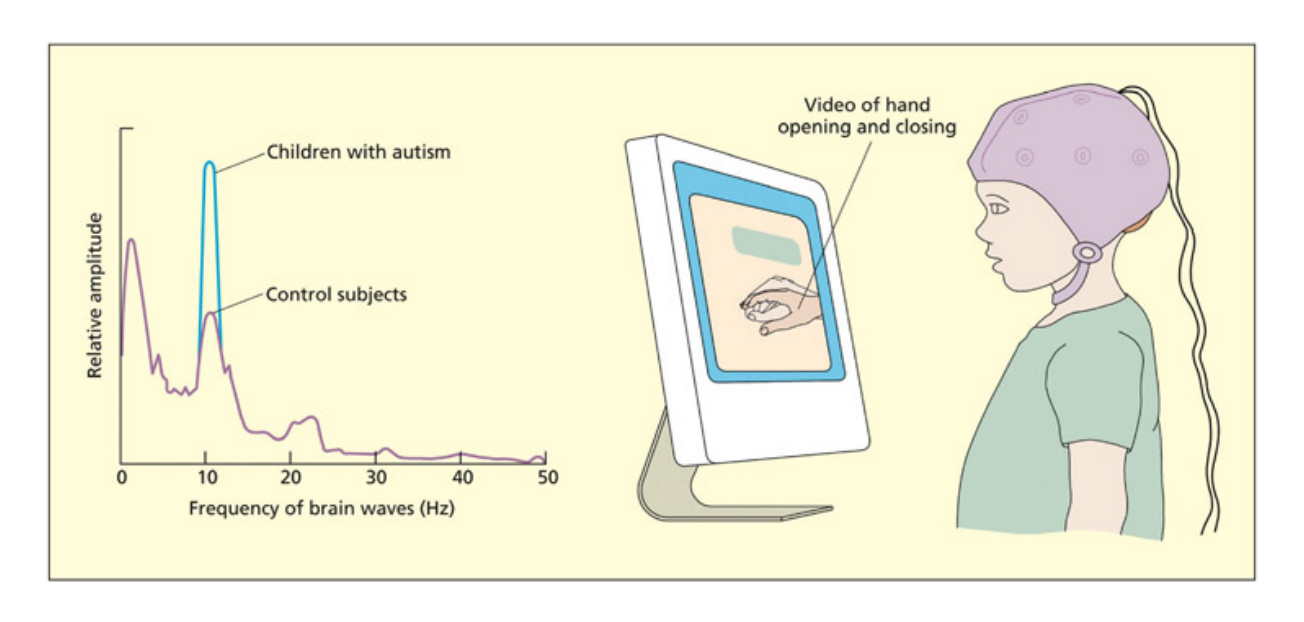
📊 그림 설명
Mu 파동(8-13 Hz)의 억제 패턴을 보여주는 EEG 데이터이다. 일반인은 자신이 행동을 수행할 때뿐 아니라 타인의 행동을 관찰할 때에도 mu 파동이 억제되며, 이는 인간의 미러 뉴런 활동 지표로 간주된다. 반면 자폐 아동은 타인의 손 동작 관찰 시 mu 억제가 현저히 감소하여, 깨진 거울 이론(broken-mirror theory)을 지지하는 증거를 제공한다.
Mu waves are EEG oscillations in the 8–13 Hz range that are reduced both when performing an action and when wating someone else perform an action (relative to rest). As su, they may provide a neural signature for human mirror neurons. Autistic ildren show less mu suppression when wating others perform a hand action, whi provides evidence in support of broken mirror theory. From Ramaandran and Oberman, 2006. Reproduced with permission from Lucy Reading-Ikkanda for Scientific American Magazine.
Temporal poles
Summary
이 섹션에서는 전두측두피질(Temporal poles)이 언어 처리와 의미 기억(semantic memory) 작업에서 활성화되는 영역임을 설명하며, Frith and Frith(2003)는 이 영역이 현재의 사회적 또는 감정적 맥락을 명확히 하기 위한 의미 단위(semas) 생성에 관여한다고 제안한다. Zahn et al.(2007)의 fMRI 연구에 따르면, 이 영역은 사회적 개념(예: 용감–영광스러운) 간의 비교에 비해 비사회적 개념(예: 영양–유용) 간의 비교보다 더 강하게 반응하는 것으로 나타났다. 또한, 이 영역을 활성화시키는 멘탈라이징(mentalizing) 작업은 언어 자극에 국한되지 않으며, 예를 들어 Castelli et al.(2000)은 서로를 격려하거나 유도하는 삼각형의 상호작용을 활용한 비언어적 자극 실험에서도 활성화가 관찰되었다. 이는 Temporal poles가 언어적 및 비언어적 사회적 맥락 해석에 모두 중요한 역할을 한다는 점을 시사한다.
is region is normally activated in tasks of language and semantic memory. Frith and Frith (2003) suggest that this region is involved with generating semas that specify the current social or emotional context, as well as in semantics more generally. Zahn et al. (2007) report an fMRI study suggesting that this region responds to comparisons between social concepts (e.g. brave–honorable) more than mated nonsocial concepts (e.g. nutritious– useful). Not all the tests of mentalizing that activated this region involved linguistic stimuli. For example, one study used triangles that appeared to interact by, say, asing or encouraging ea other (Castelli et al., 2000).
Medial prefrontal cortex
Summary
이 섹션에서는 중간 전두엽(medial prefrontal cortex)이 멘탈라이징(mentalizing)과 이론적 사고(theory-of-mind)에 핵심적인 역할을 한다는 연구 결과를 다룬다. Frith와 Frith(2003)는 이 영역이 기존의 모든 기능적 영상(functional imaging) 연구에서 사람에 대한 사고(예: 타인의 정신 상태 추론)에 특별히 반응하며, 컴퓨터나 개(기타 실체)에 대한 사고보다 더 강한 활성화를 보인다고 보고했다. 또한, 언어의 비유(예: “당신의 방은 돼지sty”)나 아이러니(예: “피터는 잘 읽는다. 셰익스피어조차 읽어봤다”)와 같은 언어의 실용적 의미(pragmatics) 처리에도 관여하며, 이와 관련된 기능 장애는 자폐 스펙트럼 장애 환자에서 관찰된다. Krueger 등(2009)은 이 영역이 행동, 주체, 목표, 물체, 신념 등 다양한 정보를 결합하여 사회적 사건(social event)을 생성하는 데 기여한다고 제안하며, 특히 자기와 유사한 타인(similar others)에 대한 판단에 더 강하게 반응하는 하위 영역이 존재한다고 설명한다. 기능적 영상 및 뇌 손상 연구에 따르면, 이론적 사고(theory-of-mind)와 관련된 주요 영역은 측두극(temporal poles), 측두-후두 접합부(temporal-parietal junction), 중간 전두엽(medial frontal lobes) 및 전두정두엽(anterior cingulate)이다. 이 영역의 기능 장애는 사회적 상호작용 및 언어 이해에 직접적인 영향을 미치며, 특히 자폐 스펙트럼 장애에서 이론적 사고 능력(theory-of-mind)의 결여와 연결된다.
Frith and Frith (2003) reported that this region is activated in all functional imaging tasks of mentalizing to that date. Functional imaging studies reliably show that this region responds more to thinking about people than thinking about other entities su as computers or dogs (e.g. Mitell et al., 2002, 2005); and thinking about the minds of people than thinking about their other aributes su as their physical aracteristics (Mitell et al., 2005), Some studies of patients with frontal lobe damage have suggested that the medial regions are necessary for theory-of-mind (Stuss et al., 2001b; Roca et al., 2011). is region also seems to be implicated in the pragmatics of language su as irony (“Peter is well read. He has even heard of Shakespeare”) and metaphor (“your room is a pigsty”; Boini et al., 1994). Interestingly, people with autism have difficulties with this aspect of language (Happe, 1995). In su instances, the speaker’s intention must be derived from the ambiguous surface properties of the words (e.g. the room is not literally a pigsty). Functional imaging suggests that this region is involved both in theory-of-mind and in establishing the pragmatic coherence between ideas/sentences, including those that do not involve mentalizing (Ferstl & von Cramon, 2002).
Krueger et al. (2009) argue that the function of this region is to bind together different kinds of information (actions, agents, goals, objects, beliefs) to create what they term a “social event.” ey note that within this region some sub-regions respond more when participants make judgments about themselves, and also judgments about others who are considered to be similar to themselves. is suggests that this region is not aributing mental states per se, but is considering the self in relation to others. e notion of creating internal social events could also explain some of the findings of the role of this region in linking ideas in story comprehension (Ferstl & von Cramon, 2002).
📊 그림 설명
마음 이론(theory-of-mind)에 관여하는 세 가지 핵심 뇌 영역을 음영으로 표시한 뇌 도식이다. 측두극(temporal poles, 좌상), 측두-두정엽 접합부(temporal-parietal junction, 우상), 내측 전두엽 및 전대상피질 일부(하단)가 포함된다. 이 영역들은 기능적 영상 연구와 뇌 손상 연구에서 멘탈라이징의 신경 기반으로 일관되게 확인되었다.
Functional imaging and lesion studies implicate three important regions in theory-of-mind (shaded): temporal poles (top le), temporal-parietal junction (top right), and the medial frontal lobes including a portion of the anterior cingulate (boom; note the anterior cingulate is drawn as divided into functionally separate areas).
From Frith and Frith, 2003. Reprinted by permission of Royal Society of London and the author.
시험 팁
Theory-of-mind의 3대 핵심 영역을 기억하는 방법: “TMP” = Temporal poles(측두극: 사회적 맥락/의미), Medial prefrontal cortex(내측전두피질: 자기-타인 관계, 사회적 사건 구성), Parietal junction(측두-두정엽 접합부: 타인의 믿음/의도 표상). 이 세 영역은 실행 기능(executive function) 네트워크나 감정 평가 네트워크와는 구별되는 별도의 네트워크를 형성한다.
Temporal-parietal junction
Summary
이 섹션에서는 후두-두정엽(temporal-parietal junction)이 멘탈라이징(mentalizing)과 생물학적 운동(perception of biological motion), 시선 방향(eye gaze), 움직이는 입, 살아 있는 물체 등에 대한 인식 과정에서 활성화되는 뇌 영역임을 강조한다. 이 영역은 타인의 정신 상태(mental states)를 이해하는 데 핵심적인 역할을 하며, 특히 가짜 믿음(false belief) 작업과 같은 이론적 사고(theory-of-mind)를 요구하는 작업에서 활성화가 관찰된다. Samson et al.(2004)의 연구에 따르면, 이 영역의 손상은 신체 인식의 어려움만으로 설명할 수 없는 이론적 사고(theory-of-mind) 작업의 실패와 관련이 있으며, Saxe and Kanwisher(2003)는 가짜 사진(false photograph) 작업과 가짜 믿음(false belief) 작업의 비교에서 이 영역이 활성화되는 것을 밝혀냈다. 또한 Saxe and Powell(2006)은 이 영역이 주관적 상태(예: 배고픔, 피로)보다 내용 있는 정신 상태(예: 생각, 믿음)의 대입(attribution)에 반응함을 보여주며, 이는 타인에 대한 사고(thinking about others)가 아닌 정신 상태(mental states)에 특화된 기능을 암시한다. 이러한 결과는 후두-두정엽이 단순한 행동/인물 인식을 넘어 정신 상태의 표현과 처리에 관여하는 신경 기초를 형성함을 시사한다.
is region tends to be not only activated in tests of mentalizing, but also in studies of the perception of biological motion, eye gaze, moving mouths, and living things in general. ese skills are clearly important for detecting other “agents” and processing their observable actions. Simulation theories argue that mentalizing need not involve anything over and above action perception. However, it is also conceivable that this region goes beyond the
processing of observable actions, and is also concerned with representing mental states. Patients with lesions in this region fail theory-of-mind tasks that can’t be accounted for by difficulties in body perception (Samson et al., 2004). Saxe and Kanwisher (2003) found activity in this region, on the right, when comparing false belief tasks (requiring mentalizing) with false photograph tasks (not requiring mentalizing but entailing a conflict with reality). e result was also found when the false photograph involved people and actions, consistent with a role in mentalizing beyond any role in action/person perception. Saxe and Powell (2006) have shown that this region responds to aribution of contentful mental states (su as thoughts and beliefs) rather than subjective states (su as hunger or tiredness). is suggests that it may have a role over and above “thinking about others” whi they claim is specific to thinking about mental states.
Evaluation
Performance on tasks of theory-of-mind typically requires the use of several nonspecialized (i.e. domain general) processes, including language processing, executive functions and action perception. e controversy lies in whether these meanisms are the only ones that are needed to account for theory-of-mind (in some forms of simulation theory) or whether there is additionally the need for a specialized (i.e. domain-specific) meanism that is specific to representing the mental states of others. Functional imaging and brain-lesion studies highlight the importance of several key regions in theory-of-mind, but the extent to whi these regions are specific to theoryof-mind is controversial. Autism still offers the most convincing evidence for a specialized meanism.
Summary and Key Points of the Chapter
Summary
이 섹션에서는 감정 모델의 현대적 관점을 정리하며, 이산적 감정(discrete emotions)과 연속적 감정 범주(continuous range of emotional processes) 간의 차이를 설명한다. 감정 처리에 관여하는 뇌 영역(예: 편도체, 인서라, 전정두정엽, 전측 기저핵)은 각각 정서적 학습(예: 두려움 조건화), 신체 관련 감정 경험, 사회적 자극 평가, 보상 예측 등에 핵심적인 역할을 수행하며, 이는 사회적 및 비사회적 자극 모두에 적용된다. 표정 인식(facial expression recognition)과 시선 감지(gaze detection)는 서로 다른 신경 메커니즘에 기반하며, 상위 측두선(superior temporal sulcus)은 시선 방향 인식에, 감각-운동 시뮬레이션(sensorimotor simulation)은 표정의 감정적 요소 인식에 중요한 기능을 한다. 공감(empathy)은 미러링(mirroring) 및 시뮬레이션 이론(simulation theory)을 통해 설명되지만, 사회적 지식과 멘탈라이징(mentalizing) 과정의 조합이 필요하다. 자폐 스펙트럼 장애(autism) 환자는 이론적 사고(theory-of-mind) 기능 장애와 미러 시스템(mirror system)의 기능 이상이 나타나며, 이는 사회적 인지 및 감정 인식의 어려움과 연관된다. 이론적 사고(theory-of-mind)와 미러 시스템(mirror system) 중 어느 것이 자폐의 핵심 결함인지에 대해서는 여전히 논쟁 중이다. 중간 전두엽(medial prefrontal cortex)과 후두-두정엽(temporoparietal junction)은 타인의 정신 상태를 이해하는 데 핵심적인 역할을 하며, 이는 도메인 특화 모듈(domain-specific module)인지에 대한 논쟁의 중심이 된다.
- Contemporary models of emotion differ with respect to whether there are a finite number of discrete emotions or whether there is a continuous range of emotional processes but with categories of emotion being constructed through interactions with nonaffective processes (e.g. appraisal, language).
- Different regions of the “emotional brain” have different functions; although these functions do not precisely map on to discrete categories of emotion (su as fear, happiness, etc.). e amygdala has a key role in emotional learning and memory (e.g. fear conditioning); the insula and anterior cingulate have key roles in bodily related aspects of emotion experience; the orbitofrontal cortex is crucial for the appraisal and control of social and emotional stimuli; and the ventral striatum has a key role in reward prediction.
- e functions of the emotional brain described above operate in both the social realm (e.g. when the stimuli consist of conspecifics) and the nonsocial realm.
- Recognizing facial expressions, processing eye gaze, and recognizing facial identity depend on different neural meanisms. e superior temporal sulcus is particularly important for gaze detection, whereas recognizing expressions depends, at least in part, on simulating the affective and sensorimotor components of that expression.
- Mirroring (and simulation theory) is an important aspect of explaining empathy, but there is more to empathy than this. Empathy is modulated be social knowledge of others and may also require a mentalizing component.
- People with autism may la a theory-of-mind meanism, but impaired theory-of-mind alone cannot explain the full paern of strengths and weaknesses in autism. It is unclear whether theory-of-mind is the core deficit in autism or whether a defective mirror system is the core deficit.
inking about the mental states of others involves a core network of regions (including temporoparietal junction and medial prefrontal cortex) that differ from those involved in executive functions more generally, or involved in emotional evaluation or person perception. Whether it is a domain-specific module remains contested.

📊 그림 설명
이 장에서 다룬 주요 개념들의 관계를 정리한 요약 도식이다. 감정 처리의 핵심 뇌 영역(편도체, 인서라, 안와전두피질, 전대상피질, 복측선조체)과 사회적 인지(얼굴 표정 인식, 시선 감지, 공감, 멘탈라이징)의 신경 기전이 종합적으로 제시되어 있다. 사회적 뇌와 감정적 뇌의 상호작용을 전체적으로 조망할 수 있는 참고 자료이다.
Visit the companion website at www.psypress/cw/ward for videos on key topics covered in this apter, references to key papers and readings, multiple oice questions and interactive glossary flashcards.
Example Essay Questions
Summary
이 섹션에서는 감정의 이산적 범주(discrete categories) 존재 여부, 편도체(amygdala)와 전정두정엽(orbitofrontal cortex)의 감정 처리 역할 차이, 자폐 스펙트럼 장애(autism)가 의식적 결여(mind-blindness)로 설명될 수 있는지, 공감(empathy)과 이론적 사고(theory-of-mind)가 시뮬레이션(simulation) 과정으로 설명될 수 있는 범위, 얼굴 정체성(facial identity), 표정 인식(expression recognition), 시선 인식(gaze recognition)의 메커니즘 차이, 그리고 미러 시스템(mirror systems)의 사회 인지(social cognition)에 대한 역할 등 6가지 주요 질문을 제시한다. 이는 이전 섹션에서 다룬 감정의 생물학적 기초, 감정 관련 뇌 영역(예: 편도체, 전정두정엽, 인서라, 전두정두엽)의 기능, 미러 뉴런(mirror neurons)과 시뮬레이션 이론(simulation theory)의 신경 기전, 자폐 스펙트럼 장애와 이론적 사고(theory-of-mind) 결손, 감정 표현(emotional expression)의 진화적 기원, 사회적 참조(social referencing) 및 정서적 적응(emotional adaptation) 등과 연계된다. 특히, 편도체는 정서적 자극(emotional stimuli)의 학습 및 두려움 조건화(fear conditioning)에 핵심 역할을 하며, 전정두정엽은 자극의 맥락적 가치(contextual value) 평가에 중대한 영향을 미친다는 점에서 두 구조의 기능 차이가 강조된다. 또한, 미러 시스템의 손상이 자폐 스펙트럼 장애의 사회 인지(social cognition) 장애와 연결되는 broken-mirror 이론(Broken-mirror theory)과 이론적 사고(theory-of-mind)의 신경 기반(예: 중간 전두엽(medial prefrontal cortex), 후두-두정엽(temporal-parietal junction))을 설명하는 데 중요한 역할을 한다. 시뮬레이션 이론(simulation theory)은 타인의 감정 상태를 이해하기 위해 자신의 감각-운동 경로(sensorimotor pathways)를 통해 미러링(mirroring)을 유도하는 메커니즘으로, 공감(empathy) 및 이론적 사고(theory-of-mind)의 핵심 요소로 작용하며, 자폐 스펙트럼 장애 환자의 미러 시스템(mirror system) 기능 이상과 관련된 mu 진동(mu oscillations)의 감소가 이에 대한 실증적 근거로 제시된다.
- Are there discrete categories of different emotions?
- Contrast the different roles of the amygdala and orbitofrontal cortex in emotion processing?
- Can autism be explained as “mind-blindness”?
- To what extent can empathy and theory-of-mind be explained by a process of simulation?
- To what extent are recognizing facial identity, expression recognition, and gaze recognition served by different meanisms?
- What is the role of mirror systems in social cognition?
Recommended Further Reading
Summary
이 섹션에서는 감정의 생물학적 기초, 뇌 기능, 사회적 인지 및 관련 질환(예: 자폐 스펙트럼 장애, Capgras 증후군)을 다루며, 감정(emotion)이 보상성(rewarding) 또는 처벌성(punishing) 자극에 반응하여 발생하는 내재적 상태로 정의된다. 주요 뇌 영역(neural substrates)으로는 편도체(amygdala), 인서라(insula), 전정두정엽(orbitofrontal cortex), 전두정두엽(anterior cingulate cortex) 등이 언급되며, 이들은 정서적 학습(예: 두려움 조건화), 사회적 자극 평가, 보상 예측 등에 핵심적인 역할을 수행한다. 또한, 미러 시스템(mirror systems)과 시뮬레이션 이론(simulation theory)은 타인의 감정을 이해하는 데 중요한 신경 기전으로, 공감(empathy) 및 이론적 사고(theory-of-mind)와 연결된다. 자폐 스펙트럼 장애(autism)는 미러 시스템(mirror system) 기능 장애와 관련된 사회적 인지(social cognition)의 이상을 설명하는 broken-mirror 이론과 연결되어 있으며, Capgras 증후군은 표정 인식 및 사회적 참조(social referencing)의 이상과 관련된 정서적 반응(emotional response) 결여를 특징으로 한다. 감정 표현(emotional expression)은 찰스 다윈의 이론에 따라 종 간 공유되는 본능적 특성으로, 공포(fear)나 분노(anger)와 같은 감정은 동물 간의 감정 상태 해석에 기능적 역할을 한다. 이 섹션은 감정 이론의 다양성과 진화적 적응 가치, 의식적(인간)과 무의식적 과정의 복합성을 강조하며, 감정은 단순한 신체 감각으로 환원될 수 없다는 점을 명확히 한다.
- Hill, E. L. & Frith, U. (2004). Autism: Mind and brain. Oxford, UK: Oxford University Press (also published as an issue of Philosophical Transactions of the Royal Society of London B (2003), 358, 277–427).
- Lane, R. D. & Nadel, L. (2000). Cognitive neuroscience of emotion. Oxford, UK: Oxford University Press. A collection of papers covering many of the issues related to the first half of this apter.
- Saxe, R. & Baron-Cohen, S. (2006). Theory of mind. Hove, UK: Psyology Press (also published as an issue of Social Neuroscience, 1, 135–416).
- Ward, J. (2012). The student’s guide to social neuroscience. New York: Psyology Press. An accessible introduction to all topics in the area. Start here.