Overview
- 연구 배경: 외부 세계의 인식과 이해를 위한 뇌 내 신경 표현의 형성 메커니즘을 밝히는 것이 핵심 목표
- 핵심 방법론:
- 단일 세포 기록을 통해 특정 자극에 대한 뉴런의 반응률을 측정하여 정보 처리 기전 분석
- EEG/ERP를 활용해 뇌의 전기적 활동을 비침습적으로 분석하고, 인지 이벤트와의 연관성을 탐구
- 주요 기여:
- 스파스 분포 인코딩(sparse distributed coding)이 특정 자극(예: 특정 인물의 얼굴)에 대한 뉴런 반응의 주요 메커니즘임을 실증
- 시간적 인코딩(temporal coding)이 공간적으로 분리된 뉴런 집단 간 정보 통합에 기여함을 보여줌
- 실험 결과:
- 인간의 중간 뇌엽에서 할리 베리와 같은 특정 인물에만 반응하는 뉴런을 확인(예: 5명의 연예인 중 특정 인물에 대한 반응률 2.3배 이상 증가)
- 시선 방향에 대한 반응에서 아래 시선(downward gaze)에 대한 뉴런 활성도가 상대적으로 40% 이상 높음
- 한계점: 단일 세포 기록은 동물 실험에 한정되며, EEG는 공간 해상도가 낮아 특정 뇌 영역 활성화를 정확히 파악하기 어려움
📋 목차
대단원 구조
- Chapter 3 The electrophysiological brain — 전기생리학적 뇌 연구의 개요 및 신경 표현 개념 소개
- 1.1 Representations — 정신적 표현과 신경적 표현의 구분
- 1.2 Electroencephalography (EEG) — EEG 기법의 기본 원리와 인지 연구에서의 활용
- In Search of Neural Representations: Single-Cell Recordings — 단일 세포 기록을 통한 신경 표현 탐구
- 2.1 How are single-cell recordings obtained? — 단일 세포 기록의 획득 방법과 기술적 한계
- 2.2 Distributed versus sparse coding — 분산 인코딩과 희소 분산 인코딩의 비교
- 2.3 Rate coding — 뉴런의 발화 빈도를 통한 정보 인코딩
- 2.4 Temporal coding — 발화 동기화를 통한 시간적 정보 인코딩
- Electroencephalography and Event-Related Potentials — EEG와 ERP의 원리 및 응용
- 3.1 How does EEG work? — EEG 신호의 생리학적 기초와 전극 배치 시스템
- 3.2 Rhythmic oscillations in the EEG signal — 알파, 베타, 감마 대역의 인지적 기능 연관성
- 3.3 Dipole — 다이폴 개념과 ERP 신호의 물리적 원리
- Some Practical Issues to Consider When Conducting EEG/ERP Research — EEG/ERP 실험의 실용적 고려사항
- 4.1 What behavioral measures should be obtained? — 행동 측정 지표의 선택 기준
- 4.2 How can interference from eye movement be avoided? — 눈 움직임 간섭의 제거 방법
- Mental Chronometry in Electrophysiology and Cognitive Psychology — 전기생리학과 인지심리학의 정신적 시간 측정
- 5.1 Additive factors method — 반응 시간을 인지 단계별로 분리하는 방법
- 5.2 Investigating face processing with ERPs and reaction times — ERP와 반응 시간을 활용한 얼굴 처리 연구
- 5.3 Endogenous and exogenous ERP components — 외생성과 내생성 ERP 성분의 구분
- 5.4 Dipole modeling — 역 문제 해결을 위한 다이폴 모델링
- 5.5 Why are Caricatures Easy to Recognize? — 카리커처의 빠른 인식 메커니즘
- 5.6 Magnetoencephalography (MEG) — MEG 기술의 원리와 EEG 대비 장점
Chapter 3 The electrophysiological brain
Summary
정신적 표현(mental representation)과 신경적 표현(neural representation)은 일대일 대응이 아니며, 뉴런의 반응 특성이 외부 세계의 특성과 상관관계를 갖는 방식으로 정보를 인코딩한다. 단일 세포 기록(single-cell recordings) 등 전기생리학적 방법을 통해 뉴런이 물리적 속성뿐 아니라 추상적 지식까지 표현할 수 있음이 밝혀졌다. 외부 세계는 뇌에 직접 복사되지 않으며, 뉴런의 반응 특성을 통해 간접적으로 표현된다.
How is it possible that the world “out there” comes to be perceived, comprehended, and acted upon by a set of neurons operating “in here”? Chapter 2 introduced some of the basic properties of the neuron, including the fact that the rate of responding of a neuron (in terms of the number of action potentials or “spikes”) is a continuous variable that reflects the informational content of that neuron. Some neurons may respond, say, when an animal is looking at an object but not when listening to a sound. Other neurons may respond when an animal is listening to a sound but not looking at an object, and still others may respond when both a sound and an object are present. As su, there is a sense in whi the world out there is reflected
by properties of the system in here. Cognitive and neural systems are sometimes said to create representations of the world. Representations need not only concern physical properties of the world (e.g. sounds, colors) but may also relate to more abstract forms of knowledge (e.g. knowledge of the beliefs of other people, factual knowledge).
Cognitive psyologists may refer to a mental representation of, say, your grandmother, being accessed in an information-processing model of face processing. However, it is important to distinguish this from its neural representation. ere is unlikely to be a one-to-one relationship between a hypothetical mental representation and the response properties of single neurons. e outside world is not copied inside the head, neither literally nor metaphorically; rather, the response properties of neurons (and brain regions) correlate with certain real-world features. As su, the relationship between a mental representation and a neural one is unlikely to be straightforward. e electrophysiological method of single-cell recordings has been used to investigate questions related to neural representations, and this method will be considered first in this apter.
Key Terms
Representations
Properties of the world that are manifested in cognitive systems (mental representation) and neural systems (neural representation).
정신적 표현은 인지 시스템에서, 신경적 표현은 신경 시스템에서 외부 세계의 속성을 반영한다.
Single-cell recordings (or single-unit recordings)
Measure the responsiveness of a neuron to a given stimulus (in terms of action potentials per second).
단일 세포 기록(single-cell recordings)은 뉴런의 행동 전위(action potentials) 발생 빈도를 측정하여 특정 자극에 대한 반응성을 정량화하는 핵심 기법이다.
Electroencephalography (EEG)
Summary
EEG는 두피 전극을 통해 뇌의 전기 신호를 측정하며, 우수한 시간 해상도로 인지 사건의 상대적 타이밍을 분석하는 데 유리하다. ERP(이벤트 관련 전위)는 특정 인지 이벤트와 두피 전압 변화를 연관시키고, 진동 기반 측정도 인지 프로세스와 연결된다. fMRI는 공간 해상도가, TMS는 시공간 해상도 모두가 우수하며, 모든 방법은 궁극적으로 인지 프로세스가 뇌에 의해 어떻게 수행되는지를 탐구한다.
Measurements of electrical signals generated by the brain through electrodes placed on different points on the scalp.
e other electrophysiological method that will be considered in this apter is electroencephalography (EEG). is is based on measurements of electrical signals generated by the brain through electrodes placed on different points on the scalp. Changes in electrical signal are conducted instantaneously to the scalp, and this method is therefore particularly useful for measuring the relative timing of cognitive events and neural activity. e method of event-related potentials (ERP) links the amount of ange in voltage at the scalp with particular cognitive events (e.g. stimulus, response). It has also become increasingly common to link the rate of ange of the EEG signal to cognitive processes (oscillation based measures) that also depend on the good temporal resolution of EEG.
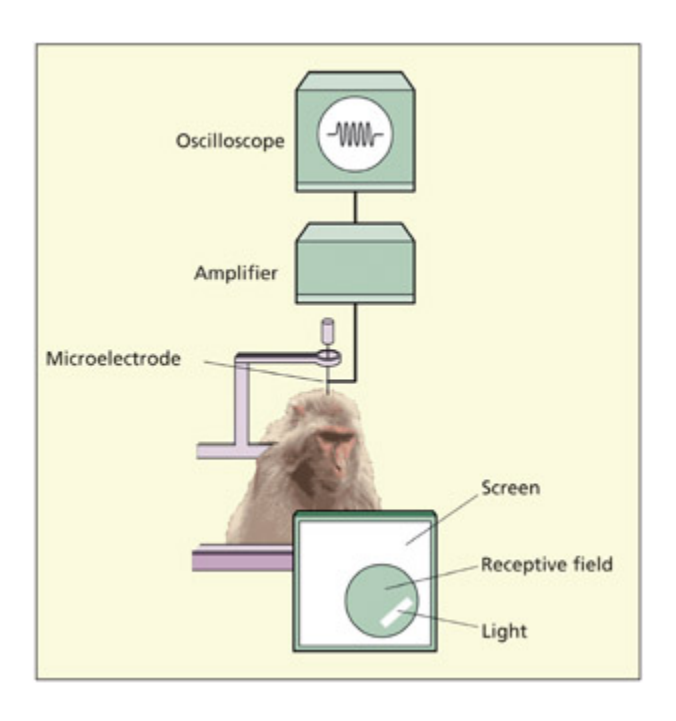
A typical experimental set-up for single-cell recording.
📊 그림 설명
단일 세포 기록의 전형적인 실험 장비 구성을 보여준다. 미세 전극이 실험 동물의 뇌에 삽입되어 특정 자극에 대한 뉴런의 행동 전위를 기록하며, 기록된 신호는 증폭기와 컴퓨터를 통해 분석된다.
ERP measurements have mu in common with the main method of cognitive psyology, namely, the reaction time measure. It is important to note that the absolute time to perform a task is not normally the thing of interest in cognitive psyology. It is of lile theoretical interest to know that one reads the word “HOUSE” within 500 ms (ms = millisecond). However, relative differences in timing can be used to make inferences about the cognitive system. For example, knowing that people are slower at reading “HoUsE” when printed in mIxEd CaSe could be used to infer that, perhaps, our mental representations of visual words are case-specific (e.g. Mayall et al., 1997). e extra processing time for “HoUsE” relative to “HOUSE” may reflect the need to transform this representation into a more standard one. Other methods in cognitive neuroscience are sensitive to measures other than timing. For example, functional imag ing methods (su as fMRI) have a beer spatial resolution than temporal resolution (see Chapter 4). Lesion methods tend to rely on meas uring error rates rather than reaction times (see Chapter 5). Methods su as transcranial magnetic stimulation (TMS) have both good spatial and temporal resolution (see Chapter 5). It is important to stress that all these methods converge on the
question of how cognitive pro cesses are carried out by the brain. Just because one method is sensitive to timing differences and another is sensitive to spatial differences does not mean that the methods just tell us when and where. e “when” and “where” constitute the data, and the “how” is the theory that accounts for them.
In Search of Neural Representations: Single-Cell Recordings
Summary
단일 세포 기록(single-cell recordings)은 침습적 방법으로 전극을 신경세포 내부 또는 외부에 삽입하여 행동 전위를 측정하며, 주로 마취된 실험 동물에서 수행된다. 세포 외 기록(extracellular recording)이 포유류에서 일반적이며, 다중 전극 배열 기술로 100개 뉴런의 동시 기록이 가능해졌다. 비침습적 두피 측정으로는 신호가 약하고 노이즈가 커서 단일 뉴런 기록이 불가능하다.
How are single-cell recordings obtained?
Summary
단일 세포 기록은 전극을 세포 내(intracellular) 또는 세포 외(extracellular)에 삽입하여 행동 전위의 발화율을 측정하는 침습적 방법이다. 포유류 뇌에서는 뉴런 크기가 작아 세포 외 기록이 일반적이며, 전신마취 하에 수행된다. 다중 세포 기록(multi-unit recordings)은 인접 뉴런의 활동을 동시에 기록한 후 알고리즘으로 개별 기여를 분리하며, 인간에서는 뇌 수술 중 드물게 수행된다.
By measuring anges in the responsiveness of a neuron to anges in a stimulus or anges in a task, it is possible to make inferences about the building blos of cognitive processing. e action potential is directly measured in the method of single-cell (and multi-unit) recordings. Singlecell recordings can be obtained by implanting a very small electrode either into the neuron itself (intracellular recording) or outside the membrane (extracellular recording) and counting the number of times that an action potential is produced (spikes per second) in response to a given stimulus (e.g. a face). is is an invasive method. As su, the procedure is normally conducted on experimental animals only. e electrodes are implanted during full anesthesia, and the recordings do not cause the animal pain. Extracellular recordings are the norm in the mammalian brain due to the small size of neurons. e method is occasionally conducted on humans under going brain surgery (see Engel et al., 2005). It is impossible to measure action potentials from a single neuron noninvasively (i.e. from the scalp) because the signal is too weak and the noise from other neurons is too high.
An electrode may pi up on activity from multiple nearby neurons and, when used in this way, is referred to as multi-cell (or multi-unit) recordings. Special algorithms can then be applied to separate the combined
signal into individual contributions from different neurons. Tenology has now advanced su that it is possible to simultaneously record from 100 neurons in multi-electrode arrays.
주의
단일 세포 기록(single-cell recording)은 침습적 방법이므로 주로 동물 실험에서만 가능하다. 두피에서 측정하는 EEG와 절대 혼동하지 말 것. 단일 세포 기록은 개별 뉴런의 발화율을 직접 측정하지만, EEG는 수백만 뉴런의 시냅스 후 전류를 합산한 신호이다.
Key Terms
Event-related potential (ERP)
The average amount of change in voltage at the scalp that are linked to the timing of particular cognitive events (e.g. stimulus, response).
ERP(이벤트 관련 전위)는 특정 인지 이벤트와 연관된 두피 전압 변화의 평균치를 측정하는 비침습적 기법이다. 단일 세포 기록과 달리 인지 시스템의 시간적 동작을 대규모로 추적할 수 있으며, 우수한 시간 해상도로 인지 프로세스의 타이밍 분석에 핵심적 도구로 활용된다.
Reaction time
The time taken between the onset of a stimulus/event and the production of a behavioral response (e.g. a button press).
반응 시간(reaction time)은 자극 시작과 행동 반응(예: 버튼 누름) 사이의 시간 간격으로, 인지 처리 속도를 평가하는 핵심 지표이다. 자극 인식, 결정, 운동 실행 등 다양한 인지 단계의 시간적 특성을 반영하며, ERP와 결합하여 신경 활동의 타이밍을 분석하는 데 활용된다.
Multi-cell recordings (or multi-unit recordings)
The electrical activity (in terms of action potentials per second) of many individually recorded neurons recorded at one or more electrodes.
다중 세포 기록(multi-cell recordings)은 하나 이상의 전극으로 여러 뉴런의 행동 전위를 동시에 측정하는 기법이다. 개별 뉴런만 분석하는 단일 세포 기록의 한계를 넘어, 신경세포 집단의 동작 패턴과 뉴런 간 상호작용을 탐구하여 신경 회로 수준의 정보 처리를 이해하는 데 기여한다.
Grandmother cell
A hypothetical neuron that just responds to one particular stimulus (e.g. the sight of one’s grandmother).
그랜드마더 셀(grandmother cell)은 특정 자극(예: 할머니의 얼굴)에만 반응하는 가설적 단일 뉴런으로, 신경적 표현의 극단적 형태이다. 현재 신경과학에서는 다수의 뉴런이 하나의 개념을 공동 인코딩하는 분산 표현(distributed representation)이 더 널리 인정받고 있으며, 이 가설은 신경 활동의 특정성과 인지 정보의 추상적 표현 간 관계를 이해하기 위한 이론적 틀로 기능한다.
Distributed versus sparse coding
Summary
시각 피질은 단순 요소에서 복잡한 형태로 통합되는 계층적 구조를 가지며, 최상위에 그랜드마더 셀 가설이 제기되었으나, Quiroga 등(2005)의 연구는 희소 분산 인코딩(sparse distributed coding)이 더 적절한 모델임을 보여주었다. 상측두구(STS) 뉴런은 시선 방향 등 사회적 단서에 특화된 반응을 보인다. 레이트 코딩(rate coding)은 발화 빈도로, 시간적 코딩(temporal coding)은 뉴런 간 동기화된 발화로 정보를 인코딩하며, Engel 등(1991)은 시각 피질에서 자극에 따른 동기화를 실증했다.
Hubel and Wiesel (1959) conducted pioneering studies of the early visual cortical areas (see Chapter 6 for detailed discussion). ey argued that visual perception is hierarical in that it starts from the most basic visual elements (e.g. small pates of light and dark) that combine into more complex elements (e.g. lines and edges), that combine into yet more complex elements (e.g. shapes). But what is the highest level of the hierary? Is there a neuron that responds to one particular stimulus? A hypothetical neuron su as this has been termed a grandmother cell because it may respond, for example, just to one’s own grandmother (Bowers, 2009). e term was originally conceived to be multi-modal, in that the neuron may respond to her voice, and the thought of her, as well as the sight of her. It is now commonly referred to as a cell that responds to the sight of her (although from any viewpoint).
Rolls and Deco (2002) distinguish between three different types of representation that may be found at the neural level:
-
- Local representation. All the information about a stimulus/event is carried in one of the neurons (as in a grandmother cell).
-
- Fully distributed representation. All the information about a stimulus/event is carried in all the neurons of a given population.
-
- Sparse distributed representation. A distributed representation in whi a small proportion of the neurons carry information about a stimulus/event.
AI 연결 — Sparse Autoencoder (SAE)와 희소 분산 표상
위의 희소 분산 표상(sparse distributed representation)은 AI에서 Sparse Autoencoder(SAE) 의 이론적 배경이 된다.
신경과학 AI (Sparse Autoencoder) 특정 자극에 소수의 뉴런만 활성화 특정 입력에 소수의 유닛/특성만 활성화 각 뉴런이 범주 내 여러 자극에 등급적 반응 각 잠재 유닛이 관련 입력에 연속적 활성도로 반응 완전 분산도, 단일 뉴런 코딩도 아닌 중간 지점 dense한 표상을 희소한 특성 공간으로 분해 특히 최근 Mechanistic Interpretability 분야에서 Anthropic 등이 LLM 내부의 dense한 활성화를 SAE로 분해하여 해석 가능한 개별 특성(feature) 을 추출하는 연구가 활발하다. 이는 “뉴런 하나 = 하나의 개념”(그랜드마더 셀)이 아니라, 소수의 해석 가능한 특성들이 조합되어 의미를 표상한다는 동일한 원리에 기반한다.

Could there be a single neuron in our brain that responds to only one stimulus, su as our grandmother? ese hypothetical cells are called “grandmother cells.”
📊 그림 설명
그랜드마더 셀 가설을 시각적으로 표현한 그림이다. 뇌 속에 특정 자극(예: 할머니)에만 반응하는 단일 뉴런이 존재할 수 있는지를 묻는 개념적 삽화로, 이 가설적 뉴런은 하나의 자극에 대해서만 선택적으로 발화한다.
Several studies have aempted to distinguish between these accounts. Bayliss et al. (1985) found that neurons in the temporal cortex of monkeys responded to several different faces (from a set of five), albeit to different degrees. Similar results have been found with mu larger sets of faces in both monkeys (Rolls & Tovee, 1995) and more recently in humans undergoing surgery for epilepsy (iroga et al., 2005). e neurons typically respond to several different stimuli from within the same category (e.g. responding to several faces but no objects). is is inconsistent with a strict definition of a grandmother cell. However, they also showed a surprising degree of specificity. In the study on humans, iroga et al. (2005) recorded from neurons in parts of the brain traditionally implicated in memory rather than perception (i.e. medial temporal lobes). ey found some neurons that responded maximally to celebrities su as Jennifer Aniston or Halle Berry, irrespective of the particular image used, clothes worn, etc. e “Halle Berry neuron” even responded to the sight of her name and to her dressed up as Catwoman, but not to other actresses dressed up as Catwoman. However, it
is impossible to conclude that the neuron only responds to Halle Berry without probing an infinite number of stimuli. ese studies speak against a fully distributed representation of personal identity and are more consistent with the notion of “sparse” coding at the top of the visual hierary.
단일 세포 측정에서의 세 가지 표상 이론 (Rolls & Deco, 2002)
1. 국소 표상 (Local Representation)
하나의 자극/사건에 대한 모든 정보가 단일 뉴런 하나에 담긴다는 입장이다. 그랜드마더 셀 가설이 대표적 사례로, 예컨대 “할머니 뉴런”은 할머니의 시각·청각·개념 정보를 혼자 인코딩한다. 그러나 Bayliss et al. (1985)에서 측두 피질 뉴런이 하나의 얼굴이 아닌 여러 얼굴에 등급적으로 반응한다는 결과가 나왔고, Quiroga et al. (2005)의 “Halle Berry 뉴런”도 Halle Berry 외 다른 자극에도 반응할 가능성을 배제할 수 없어 엄격한 국소 표상은 지지되지 않는다.2. 완전 분산 표상 (Fully Distributed Representation)
하나의 자극/사건에 대한 정보가 주어진 뉴런 집단의 모든 뉴런에 분산되어 있다는 입장이다. 그러나 Bayliss et al. (1985)에서 측두 피질 뉴런들은 얼굴에는 반응하지만 물체에는 전혀 반응하지 않았다. 모든 뉴런이 모든 자극을 코딩한다면 물체에도 반응해야 하므로, 완전 분산 표상 역시 기각된다.3. 성긴 분산 표상 (Sparse Distributed Representation) ✅
하나의 자극/사건에 대한 정보가 뉴런 집단 중 소수의 뉴런에만 분산되어 있다는 입장이다. 각 뉴런은 같은 범주 내 여러 자극에 차등적(graded)으로 반응하되, 관련 없는 범주에는 반응하지 않는다. Bayliss et al. (1985), Rolls & Tovee (1995), Quiroga et al. (2005)의 결과가 모두 이 모델과 일치하며, 현재 가장 널리 지지되는 표상 이론이다.
결론: 실험적 증거는 국소 표상(너무 극단적)과 완전 분산 표상(너무 광범위) 모두를 기각하고, 그 중간인 성긴 분산 표상을 지지한다. 시각 계층 최상위에서 소수의 뉴런이 범주 내 자극을 공동으로, 그러나 차등적으로 인코딩하는 방식이 뇌의 실제 표상 메커니즘에 가장 부합한다.
완전 분산 표상 vs 성긴 분산 표상 — 정확히 뭐가 다른가?
두 이론 모두 “여러 뉴런이 함께 정보를 표상한다”는 점에서 분산(distributed) 이라는 공통점이 있다. 차이는 “주어진 집단(given population) 중 얼마나 많은 뉴런이 참여하는가” 에 있다.
“주어진 뉴런 집단”이란?
특정 기능을 담당하는 뇌 영역의 뉴런 그룹을 말한다. 예를 들어, 측두 피질에서 시각 자극을 처리하는 뉴런 수백~수천 개가 하나의 “집단”이다.
완전 분산 표상: 할머니 얼굴을 보면, 이 집단의 모든 뉴런이 반응한다. 아버지 얼굴을 봐도, 강아지를 봐도, 컵을 봐도 — 모든 뉴런이 항상 반응한다. 자극마다 달라지는 것은 각 뉴런의 반응 패턴(어떤 뉴런이 강하게, 어떤 뉴런이 약하게)뿐이다.
성긴 분산 표상: 할머니 얼굴을 보면, 이 집단 중 소수의 뉴런만 반응한다. 나머지 대다수는 침묵한다. 아버지 얼굴을 보면 다른 소수의 뉴런이 반응한다. 각 자극마다 활성화되는 뉴런의 부분집합이 다르다.
비교 항목 완전 분산 표상 성긴 분산 표상 자극당 반응 뉴런 수 집단의 전부 집단의 소수 침묵하는 뉴런 없음 (모두 반응) 대부분 침묵 자극 구별 방식 모든 뉴런의 반응 강도 패턴 차이 어떤 뉴런이 활성화되는지 자체가 다름 범주 선택성 없음 — 얼굴이든 물체든 모두 반응 있음 — 얼굴에만 반응, 물체에는 침묵 Bayliss et al. (1985) 실험으로 보는 차이:
- 만약 완전 분산이라면 → 뉴런 a, b, c, d가 얼굴(A–E)에도 물체(F–J)에도 모두 반응해야 함 (강도 패턴만 다르게)
- 실제 결과 → 뉴런들이 얼굴에만 반응하고 물체에는 전혀 반응하지 않음 → 완전 분산 기각
- 동시에 각 뉴런이 하나의 얼굴이 아닌 여러 얼굴에 등급적으로 반응 → 국소 표상도 기각
- 결론: 소수의 뉴런이 범주 내에서만 선택적으로 반응 → 성긴 분산 표상
Some neurons code for other aspects of a stimulus than facial identity. For example, consider the paern of responding of a particular neuron taken from the superior temporal sulcus (STS) of an alert macaque monkey (Perre et al., 1992). e activity of the neuron when shown four different views of faces is compared with spontaneous activity in whi no face is shown. e neuron responds strongly to a downward gaze, both with the eyes and the whole head, but not an upward or straight-ahead gaze. In this instance, the two stimuli that elicit the strongest response (head down and head forward with eyes down) do not resemble
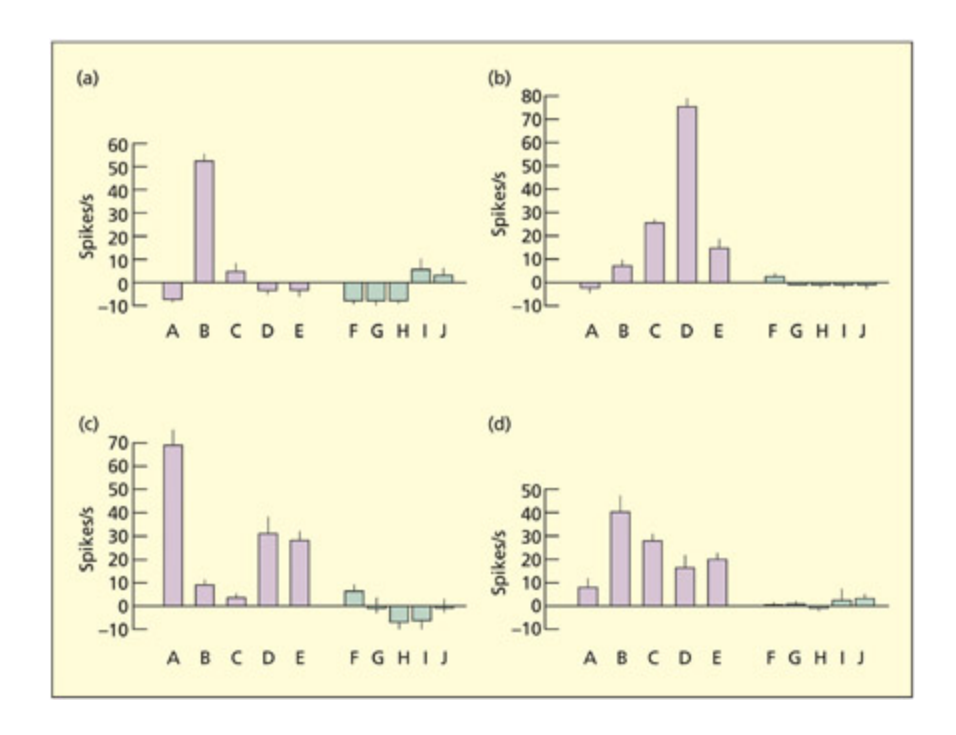
Four neurons (a, b, c, d) respond to different faces (A–E), but not different objects (F–J). ey typically respond to several faces, albeit in a graded fashion.
Reprinted from Bayliss et al., 1985. © 1985, with permission from Elsevier.
📊 그림 설명
4개의 뉴런(a, b, c, d)이 서로 다른 얼굴(A-E)에 대해 각기 다른 정도로 반응하지만, 물체(F-J)에는 반응하지 않는 패턴을 보여준다. 이는 뉴런이 범주 내 여러 자극에 등급적으로 반응하는 희소 분산 인코딩을 지지하는 증거이다.
실험 상세 — Bayliss et al. (1985)
목적: 측두 피질의 개별 뉴런이 특정 자극에 어떻게 반응하는지 확인하여, 그랜드마더 셀 가설의 타당성을 검증한다.
방법: 원숭이(macaque)의 측두 피질(temporal cortex)에 미세전극을 삽입하여 단일 뉴런의 발화를 기록했다. 자극으로 5개의 얼굴(A–E)과 5개의 물체(F–J)를 제시하고, 각 뉴런의 반응(발화율)을 비교했다.
결과:
- 4개 뉴런(a, b, c, d) 모두 얼굴에는 반응하되 물체에는 반응하지 않음 → 범주 선택성(category selectivity)
- 각 뉴런이 하나의 얼굴에만 반응한 것이 아니라, 여러 얼굴에 등급적(graded)으로 반응 → 특정 얼굴에 강하게, 다른 얼굴에 약하게
- 뉴런마다 가장 강하게 반응하는 얼굴이 다름
의의:
- 그랜드마더 셀 가설 반박: 하나의 자극에만 반응하는 뉴런(local representation)이 아님
- 완전 분산 표상도 아님: 물체에는 전혀 반응하지 않으므로, 모든 뉴런이 모든 자극을 코딩하는 것도 아님
- 희소 분산 인코딩(sparse distributed coding) 지지: 소수의 뉴런이 범주 내 여러 자극에 차등적으로 반응하는 방식으로 정보를 표상
이후 Rolls & Tovee (1995)가 더 큰 얼굴 세트로, Quiroga et al. (2005)가 인간 간질 수술 환자에서 유사한 결과를 재현하였다.
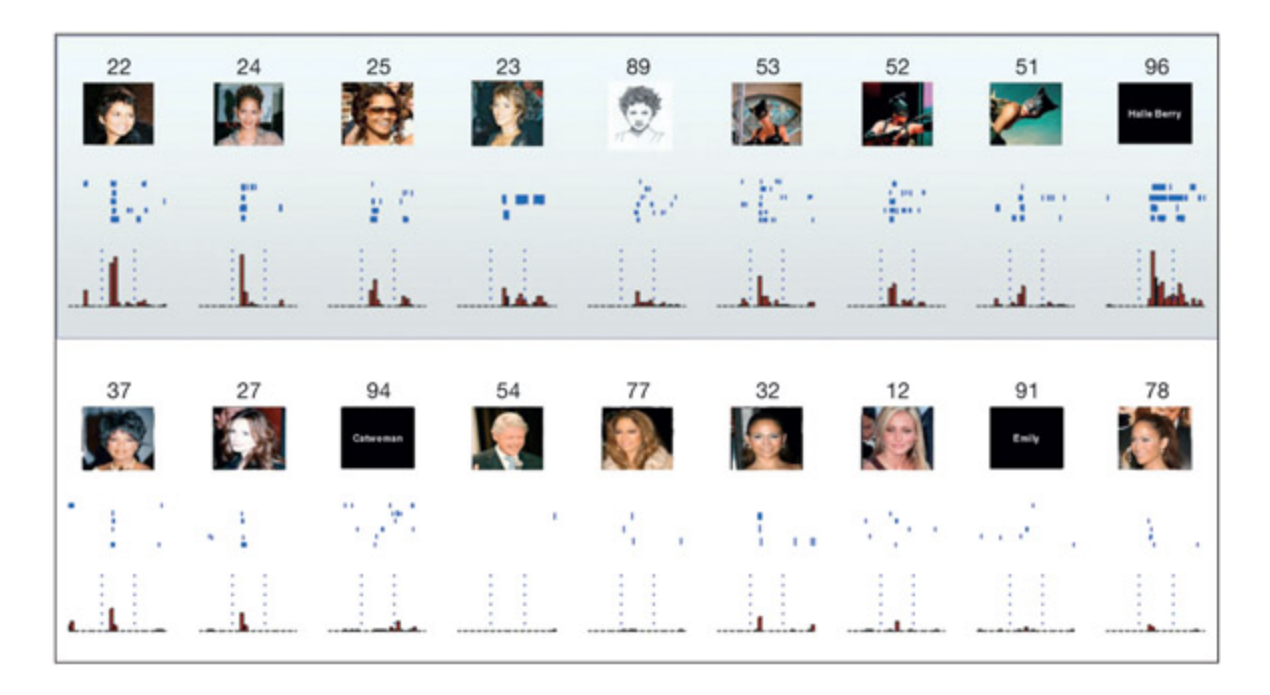
is neuron, recorded in the human medial temporal lobe, responds to Halle Berry (top panel) more than comparable stimuli (boom panel). e response of the neuron is depicted in two ways. A raster plot (blue) depicts the firing of the neuron over time (represented le-to-right horizontally) by shading in when the neuron fires. Ea row is a different recording with that stimulus. e histogram (red) sums together the number of times that the neuron fired at ea time point. From iroga et al., 2005.
📊 그림 설명
인간 내측두엽에서 기록된 뉴런이 할리 베리의 다양한 이미지(상단)에 강하게 반응하지만 유사한 다른 자극(하단)에는 반응하지 않는 패턴을 보여준다. 래스터 플롯(파란색)은 시간에 따른 개별 발화를, 히스토그램(빨간색)은 시점별 발화 빈도의 합계를 나타낸다.
실험 상세 — Quiroga et al. (2005)
목적: 인간 뇌의 개별 뉴런이 특정 인물에 대해 얼마나 선택적으로 반응하는지 조사하여, 국소 표상(그랜드마더 셀)과 분산 표상 중 어떤 모델이 더 적합한지 검증한다.
방법: 간질 수술을 위해 전극이 삽입된 인간 환자를 대상으로, 지각이 아닌 기억과 관련된 영역인 내측두엽(medial temporal lobe)에서 단일 뉴런의 활동을 기록했다. 다양한 유명인의 사진, 이름 텍스트, 변장한 모습 등 여러 형태의 자극을 제시했다.
결과:
- 특정 뉴런이 Halle Berry에 최대 반응을 보였으며, 사용된 특정 이미지, 의상 등과 무관하게 반응함
- 해당 “Halle Berry 뉴런”은 그녀의 이름 텍스트에도 반응하고, **캣우먼 복장**을 한 모습에도 반응했으나, 다른 배우가 캣우먼 복장을 입은 경우에는 반응하지 않음
- Jennifer Aniston 등 다른 유명인에 대해서도 유사한 선택적 뉴런이 발견됨
의의:
- 뉴런이 특정 시각적 특징이 아니라 추상적 인물 정체성(identity)을 인코딩함을 시사
- 그러나 무한한 자극을 모두 테스트할 수 없으므로, 해당 뉴런이 오직 Halle Berry에만 반응한다고 결론 내릴 수는 없음 → 엄격한 그랜드마더 셀 가설은 지지되지 않음
- 완전 분산 표상에도 반함: 뉴런이 모든 자극이 아닌 특정 인물에 고도로 선택적이므로
- 시각 계층 최상위에서의 성긴 분산 인코딩(sparse coding)이 가장 적합한 모델임을 지지
ea other physically, although they are related con ceptually. Coding of gaze direction may be import ant for cognitive processes involved in interpreting social cues (eye contact is perceived as a threat by many species), or for orienting aention and action systems. Perhaps there is something interesting down there that would warrant our aention.
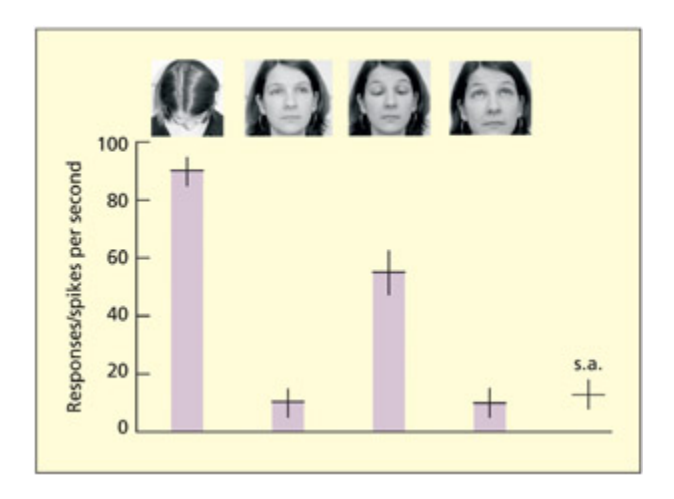
is neuron responds when gaze is oriented downwards. e activity of the neuron (spikes per second) is shown when presented with four faces and during spontaneous activity (s.a.). Adapted from Perre et al., 1992.
📊 그림 설명
상측두구(STS)의 뉴런이 시선이 아래를 향할 때 강하게 반응하는 패턴을 보여준다. 4가지 얼굴 자극과 자발적 활동(s.a.)을 비교하여, 눈이나 머리가 아래를 향하는 자극에서 발화율이 현저히 증가함을 나타낸다.
STS = Superior Temporal Sulcus(상측두고랑/상측두구). 인지신경과학에서 STS라고 하면 일반적으로 posterior STS(pSTS), 즉 상측두구의 후방 영역을 가리키는 경우가 많다. 이 영역은 시선 방향, 표정, 생물학적 움직임 등 사회적 단서를 처리하는 핵심 영역으로, 본 실험에서 시선 방향에 선택적으로 반응한 뉴런이 기록된 곳이기도 하다.
실험 상세 — Perrett et al. (1992)
목적: 상측두구(STS) 뉴런이 얼굴의 어떤 속성에 선택적으로 반응하는지, 특히 시선 방향이라는 사회적 단서에 특화된 인코딩이 존재하는지 조사한다.
방법: 각성 상태의 마카크 원숭이(macaque)의 상측두구(STS)에 미세전극을 삽입하여 단일 뉴런의 활동을 기록했다. 자극으로 4가지 서로 다른 얼굴 방향(머리 위, 머리 아래, 눈만 아래, 정면 응시)을 제시하고, 자극 없는 자발적 활동(spontaneous activity)과 비교했다.
결과:
- 해당 뉴런은 아래를 향한 시선에 강하게 반응: 머리가 아래로 향한 경우와, 머리는 정면이지만 눈만 아래로 향한 경우 모두에서 높은 발화율을 보임
- 위를 향하거나 정면을 응시하는 자극에는 거의 반응하지 않음
- 흥미롭게도, 가장 강한 반응을 이끈 두 자극(머리 아래 / 머리 정면+눈 아래)은 물리적 외양이 매우 다름에도 불구하고, “시선이 아래를 향한다”는 개념적 속성을 공유
의의:
- STS 뉴런은 단순한 시각적 특징이 아니라 시선 방향이라는 추상적·사회적 단서를 인코딩함을 시사
- 시선 방향 코딩은 사회적 상호작용 해석(눈 맞춤은 많은 종에서 위협으로 인식), 주의 방향 파악, 행동 시스템 조절 등에 기여할 수 있음
- 얼굴 정체성을 코딩하는 뉴런(Bayliss, Quiroga 등)과 달리, 얼굴의 사회적 속성을 코딩하는 별도의 신경 메커니즘이 존재함을 보여줌
e studies described above can all be classified as rate coding of information by neurons in that a given stimulus/event is associated with an increase in the rate of neural firing. An alternative way for neurons to represent information about stimuli/events is in terms of temporal coding, in that a given stimulus/event is associated with greater synronization of firing across different neurons. Engel et al. (1991) obtained multi-cell recordings from neurons in the primary visual cortex. is region contains a spatial map of the retinal image (see Chapter 6). If two regions were stimulated with a single bar of light, the two regions synronized their neural firing. But, if the two regions were stimulated by two different bars of light, there was no synronization even though both regions showed a response in terms of increased rate of firing. Temporal coding may be one meanism for integrating information across spatially separated populations of neurons.
뉴런의 두 가지 정보 인코딩 방식: Rate Coding vs Temporal Coding
1. 레이트 코딩 (Rate Coding)
특정 자극/사건이 제시될 때 뉴런의 발화 빈도(초당 행동 전위 수)가 증가하는 방식으로 정보를 표상한다. 앞서 다룬 단일 세포 기록 연구들(Bayliss et al., Quiroga et al., Perrett et al.)은 모두 레이트 코딩에 해당한다 — 특정 자극에 대해 뉴런의 발화율이 올라가는 것을 측정한 것이다.
- 측정 방법: 단일 세포 기록(single-cell recording)으로 개별 뉴런의 발화율을 정량화
- 정보 전달 원리: 발화율이 높을수록 해당 자극에 대한 반응이 강함
- 한계: 발화율만으로는 여러 자극이 동시에 존재할 때 어떤 자극들이 하나의 대상으로 묶이는지(binding) 설명하기 어려움
2. 시간적 코딩 (Temporal Coding)
특정 자극/사건이 제시될 때 서로 다른 뉴런들의 발화가 동기화(synchronization)되는 방식으로 정보를 표상한다. 발화율 자체가 아니라 뉴런 간 발화 타이밍의 일치가 핵심이다.
- 측정 방법: 다중 세포 기록(multi-cell recording)으로 여러 뉴런의 동시적 발화 패턴을 분석
- 정보 전달 원리: 같은 자극을 처리하는 뉴런들이 동기화하여 발화함으로써, 공간적으로 분리된 뉴런 집단 간 정보를 통합(binding)
- 핵심 근거 — Engel et al. (1991): 일차 시각 피질(V1)에서 다중 세포 기록을 실시한 결과, 하나의 빛 막대로 두 영역을 자극하면 두 영역의 뉴런이 발화를 동기화했지만, 두 개의 다른 빛 막대로 각각 자극하면 두 영역 모두 발화율은 증가하되 동기화는 일어나지 않았다. 즉 발화율(rate coding)만으로는 구별할 수 없는 “같은 대상인가 다른 대상인가”라는 정보가 동기화(temporal coding)를 통해 표상된다.
비교 항목 레이트 코딩 시간적 코딩 정보 단위 초당 발화 수 (firing rate) 뉴런 간 발화 동기화 (synchrony) 측정 방법 단일 세포 기록 다중 세포 기록 핵심 기능 자극의 강도·정체성 표상 공간적으로 분리된 정보의 통합 (binding) 대표 연구 Bayliss et al., Quiroga et al. Engel et al. (1991)
Evaluation
Summary
뉴런은 반응률(rate coding)과 발화 동기화(temporal coding)라는 두 가지 방식으로 정보를 인코딩하며, 이를 단일 세포 기록 및 다중 세포 기록으로 측정한다. 정보 표현은 희소 분산 인코딩(sparse distributed coding)에 기반하여 여러 뉴런의 활동이 하나의 자극을 표현하며, 이는 에너지 효율성과 고용량 기억 저장을 가능하게 한다. 분산 표현은 시냅스나 뉴런 손실 시에도 정보를 보존하고, 새로운 자극에 대한 일반화와 분류를 용이하게 한다.
Information is represented in neurons by the response rates to a given stimulus or event and, in some circumstances, by the synronization of
their firing. is can be experimentally measured by the methods of singlecell and multi-cell recordings. Both ways of representing information may depend on sparse distributed coding su that activity in several neurons is required to represent a stimulus (e.g. a particular face). e sparseness of coding conserves energy and may enable the brain to have a high memory capacity. Distributed representation may protect against information loss if synapses or neurons are lost. It may also allow the cognitive system to generalize and categorize (e.g. a novel stimulus that resembles a stored representation would partially activate this representation).
Key Terms
Rate coding
The informational content of a neuron may be related to the number of action potentials per second.
레이트 코딩(rate coding)은 뉴런의 정보 내용이 초당 행동 전위(action potentials)의 수와 관련되는 인코딩 방식이다. 시간적 코딩(temporal coding)과 대비되며, 단일 세포 기록에서 뉴런 활동을 정량화하는 기본 단위로 활용된다.
Temporal coding
The synchrony of firing may be used by a population of neurons to code the same stimulus or event.
시간적 코딩(temporal coding)은 뉴런 집단이 발화의 동기화(synchrony of firing)를 통해 동일한 자극을 인코딩하는 방식이다. 레이트 코딩과 대비되며, 공간적으로 분리된 뉴런 집단 간 정보를 통합하는 메커니즘으로 기능할 수 있다. 다중 세포 기록을 통해 동기화 패턴을 분석함으로써 신경적 표현의 시간적 차원을 탐구한다.
시험 팁
세 가지 인코딩 방식을 구분하자: 레이트 코딩(rate coding) = 발화 빈도로 정보 전달, 시간적 코딩(temporal coding) = 뉴런 간 동기화로 정보 통합, 희소 분산 코딩(sparse distributed coding) = 소수의 뉴런이 하나의 자극을 공동 표현. “그랜드마더 셀”은 극단적 국소 표현의 가설일 뿐, 실제로는 희소 분산 인코딩이 더 타당하다.
Electroencephalography and Event-Related Potentials
Summary
EEG는 두피 전극으로 뇌의 전기 신호를 측정하며, ERP는 특정 인지 이벤트와 연관된 전압 변화의 평균치를 분석한다. 반응 시간 측정보다 신경 활동의 미세한 시간적 변화를 더 정밀하게 포착할 수 있으며, 현대 인지 신경과학에서 널리 활용된다.
is section considers the basic principles behind the electrophysiological method known as electroencephalography (EEG). e following sections go on to consider some concrete examples of how EEG is used in contemporary cognitive neuroscience and contrast it with other methods used in cognitive psyology and cognitive neuroscience (principally the reaction-time measure).
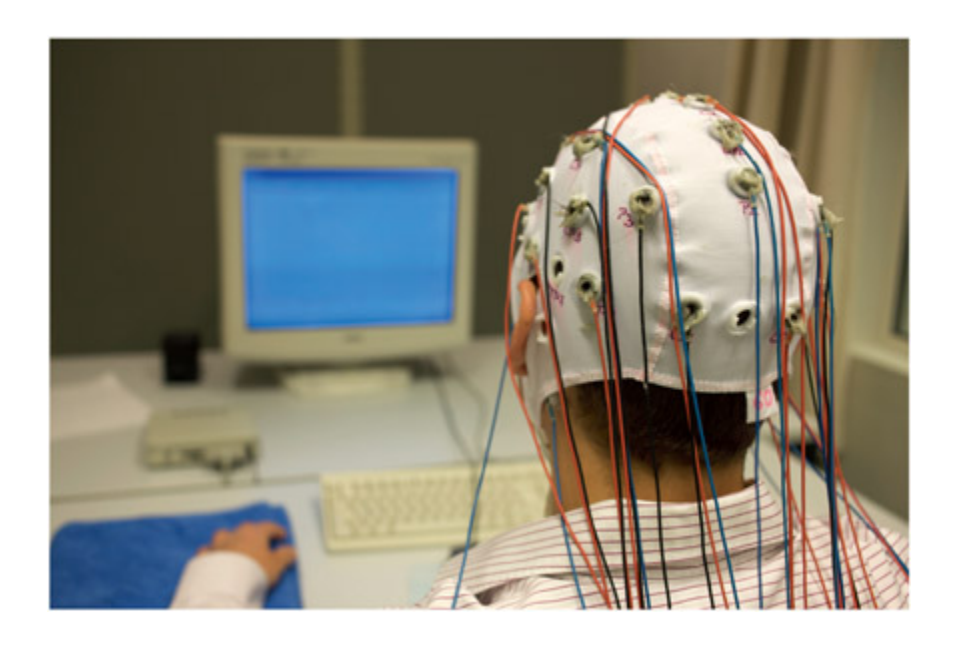
A participant in an EEG experiment. AJ Photo / HOP AMERICAIN / Science Photo Library.
📊 그림 설명
EEG 실험에 참가하는 피험자의 모습을 보여준다. 두피에 다수의 전극이 부착된 전극 캡을 착용하고 있으며, 각 전극은 뇌의 전기적 활동을 비침습적으로 기록하기 위해 특정 위치에 배치된다.
How does EEG work?
Summary
EEG 신호는 시냅스 후 수상돌기 전류(postsynaptic dendritic currents)에서 기원하며, 뉴런의 동기화된 활동과 병렬 배열이 두피에서 감지 가능한 전기장 생성에 필수적이다. 대뇌피질에서는 이 조건이 충족되지만 시상(thalamus) 등은 감지가 어렵다. 10-20 전극 시스템은 F(전두), P(두정), O(후두), T(측두), C(중심) 위치와 반구(홀수=좌, 짝수=우, z=중선)로 전극을 표기하며, 특정 위치의 신호가 원거리 활동의 영향을 받을 수 있어 EEG/ERP의 공간 해상도는 제한적이다.
e physiological basis of the EEG signal originates in the postsynaptic dendritic currents rather than the axonal currents associated with the action potential (Nunez, 1981). ese were described as passive and active currents, respectively, in Chapter 2. Electroencephalography (EEG) records electrical signals generated by the brain through electrodes placed on different points on the scalp. As the procedure is non-invasive and involves recording (not stimulation), it is completely harmless as a method. For an electrical signal to be detectable at the scalp a number of basic requirements need to be met
in terms of underlying neural firing. First, a whole population of neurons must be active in synrony to generate a large enough electrical field. Second, this population of neurons must be aligned in a parallel orientation so that they summate rather than cancel out. Fortunately, neurons are arranged in this way in the cerebral cortex. However, the same cannot necessarily be said about all regions of the brain. For example, the orientation of neurons in the thalamus renders its activity invisible to this recording method.
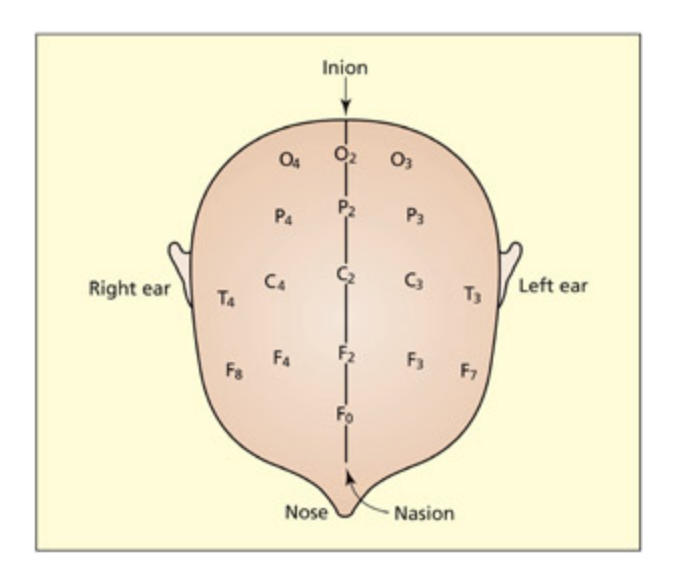
e 10–20 system of electrodes used in a typical EEG/ERP experiment.
📊 그림 설명
EEG/ERP 실험에서 사용되는 10-20 전극 배치 시스템을 도식화한 그림이다. 전극은 F(전두엽), P(두정엽), O(후두엽), T(측두엽), C(중심엽) 등 뇌 영역에 따라 배치되며, 홀수(좌반구), 짝수(우반구), z(중선)로 구분된다.
To gain an EEG measure one needs to compare the voltage between two or more different sites. A reference site is oen osen that is likely to be relatively uninfluenced by the variable under investigation. One common refer ence point is the mastoid bone behind the ears or a nasal reference; another alternative is to refer ence to the average of all electrodes. e experimental electrodes themselves are oen arranged at various locations on the scalp and oen described with reference to the so-called 10–20 system of Jasper (1958). e electrodes are labeled according to their location (F = frontal, P = parietal, O = occipital, T = temporal, C = central) and the hemisphere involved (odd numbers for le, even numbers for right, and “z” for the midline). For example, the O2 electrode is located over the right
occipital lobe, and the Fz electrode is located over the midline of the frontal lobes. It is important to stress that the activity recorded at ea location cannot necessarily be aributed to neural activity near to that region. Electrical activity in one location can be detected at distant locations. In general, EEG/ERP is not best equipped for detecting the location of neural activity (see later for further discussion).
주의
EEG와 ERP를 혼동하는 학생이 매우 많다. EEG = 연속적 뇌파 기록(raw signal), ERP = 자극에 동기화하여 여러 시행을 평균화한 결과이다. ERP는 EEG에서 추출하는 것이지, 별도의 장비로 측정하는 것이 아니다. 또한 EEG 신호의 원천은 축삭의 활동전위가 아니라 수상돌기의 시냅스 후 전류라는 점도 자주 출제된다.
Rhythmic oscillations in the EEG signal
Summary
EEG 신호의 주기적 진동은 인지 기능과 연관되며, 알파 대역(7-14 Hz) 증가는 무관한 정보 필터링, 감마 대역(30 Hz+) 증가는 물체 인식의 지각적 통합(binding)과 관련된다. ERP는 자극 제시에 대한 EEG 신호를 평균화하여 P300, N400 등의 정점으로 인지 사건의 시간적 특성을 분석한다. EEG의 빠른 시간 해상도는 fMRI와 달리 뉴런 간 동기화의 인지적 역할을 탐구하는 데 유리하다.
e EEG signal, when observed over a sufficiently long timescale, has a wave-like structure. e EEG signal tends to oscillate at different rates (also called frequency bands) that are named aer leers of the Greek alphabet: thus alpha waves reflect oscillations in the 7 to 14 Hz range, beta in the 15 to 30 Hz range, and gamma in the 30 Hz and above range (and so on). ese oscillations arise because large groups of neurons tend to be in temporal synrony with ea other in terms of their firing (action potentials) and in terms of their slower dendritic potentials (whi forms the basis of the EEG signal). It has long been established that different rates of oscillation aracterize different phases of the sleep-wake cycle (for the detailed meanisms see McCormi & Bal, 1997).
In recent decades, aempts have been made to link the relative amount of oscillations (the “power”) in different bands to different kinds of cognitive function during normal wakefulness (Ward, 2003). is section will provide only a few examples from the literature to illustrate the general principle. For instance, increases in the alpha band have been linked to increased aention. More specifically, it has been linked to filtering out of irrelevant information. If participants are asked to ignore a region of space in whi an irrelevant stimulus will later appear (a so-called distractor) then increases in the alpha band are found over electrode sites that represent that region of space (Worden et al., 2000). Alpha is also greater when aending to an internally generated image in whi external visual input is unaended (Cooper et al., 2003). An “increase in the alpha band” means that neurons
become more synronized in their electrical activity specifically in the 7 to 14 Hz range. What is less clear is why this particular neural coding should be linked to this kind of cognitive meanism rather than anges in any other frequency band.
By contrast, increases in the gamma band have been linked to perceptual integration of parts into wholes. is kind of meanism is important for object recognition (e.g. deciding that a handle and hollowed cylinder is a single object— a mug), and the general process is referred to as binding or grouping. Rodriguez et al. (1999) presented participants with an ambiguous visual stimulus that could be perceived either as a face (parts bound into a whole) or a meaningless visual paern (collection of separate parts). ey found that increased gamma synronization was linked to the face percept (Rodriguez et al., 1999).
Although gamma and delta have been linked to rather different functions (similarly for other frequency bands), it is inconceivable that there will be a one-to-one mapping between particular frequency bands and particular cognitive functions. Synronization (or desynronization) of alpha, gamma, and so on are linked to a wide range of cognitive functions and may come from different regions in the brain. However, it provides another tool within whi to understand the different meanisms that comprise cognition. Perhaps, most importantly, it suggests that there is more to cognition than the amount of brain “activity” (the standard interpretation of fMRI data) and suggests that the synchronization of brain activity (measurable in EEG because of its fast temporal resolution) has particular roles to play in cognition.
Event-related potentials (ERPs)
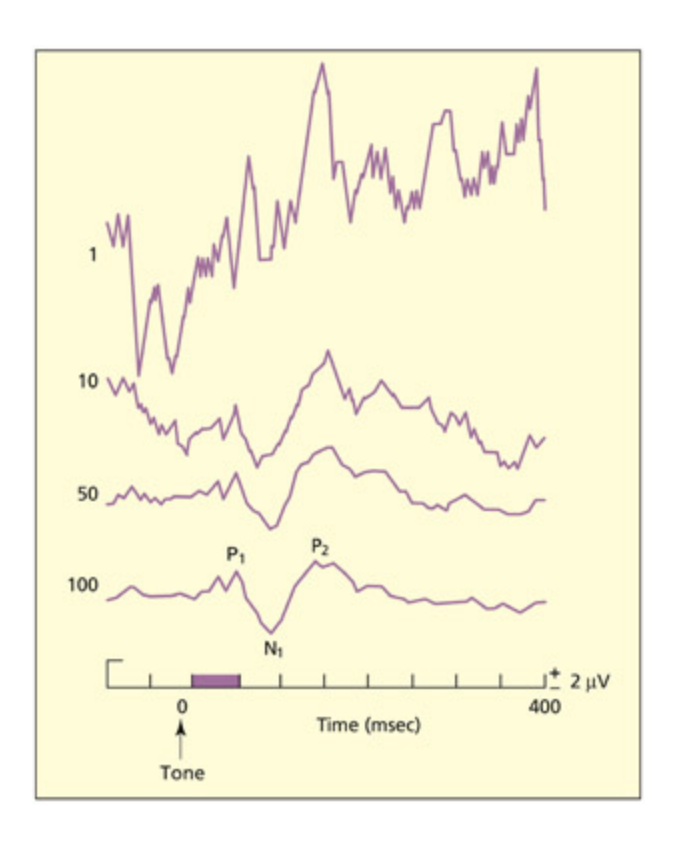
When different EEG waves are averaged relative to presentation of a stimulus (e.g. a tone), the signalto-noise ratio is enhanced and an event-related potential is observed. e figure shows the mean EEG signal to 1, 10, 50, and 100 trials.
From Kolb and Whishaw, 2002. © 2002 by Worth Publishers. Used with permission.
📊 그림 설명
자극(예: 소리) 제시에 대한 EEG 파형을 1, 10, 50, 100회 시행으로 평균화한 결과를 보여준다. 시행 수가 증가할수록 신호 대 잡음비가 향상되어 이벤트 관련 전위(ERP)의 특징적인 정점과 골이 명확해진다.
e most common use of EEG in cognitive neuroscience is not in measurements of neuronal oscillations, but rather in the method known as ERP or event-related potentials. e EEG waveform reflects neural activity from all parts of the brain. Some of this activity may specifically relate to the current task (e.g. reading, listening, calculating), but most of it will relate to spontaneous activity of other neurons that do not directly contribute to the task. As su, the signal-to-noise ratio in a single trial of EEG is very low (the signal being the electrical response to the event and the noise being the baground level of electrical activity). e ratio can be increased by averaging the EEG signal over many presentations of the stimulus (e.g. 50– 100 trials), relative to the onset of a stimulus. In general, the baground oscillatory activity (alpha, beta, etc.) will not be synronised with the onset of events and so these fluctuations are also averaged out. e results are
represented graphically by ploing time (milliseconds) on the x-axis and electrode potential (microvolts) on the y-axis. e graph consists of a series of positive and negative peaks, with an asymptote at 0 µV. is is done for ea electrode, and ea will have a slightly different profile. e positive and negative peaks are labeled with “P” or “N” and their corresponding number. us, P1, P2, and P3 refer to the first, second, and third positive peaks, respectively. Alternatively, they can be labeled with “P” or “N” and the approximate timing of the peak. us, P300 and N400 refer to a positive peak at 300 ms and a negative peak at 400 ms (not the 300th positive and 400th negative peak!).
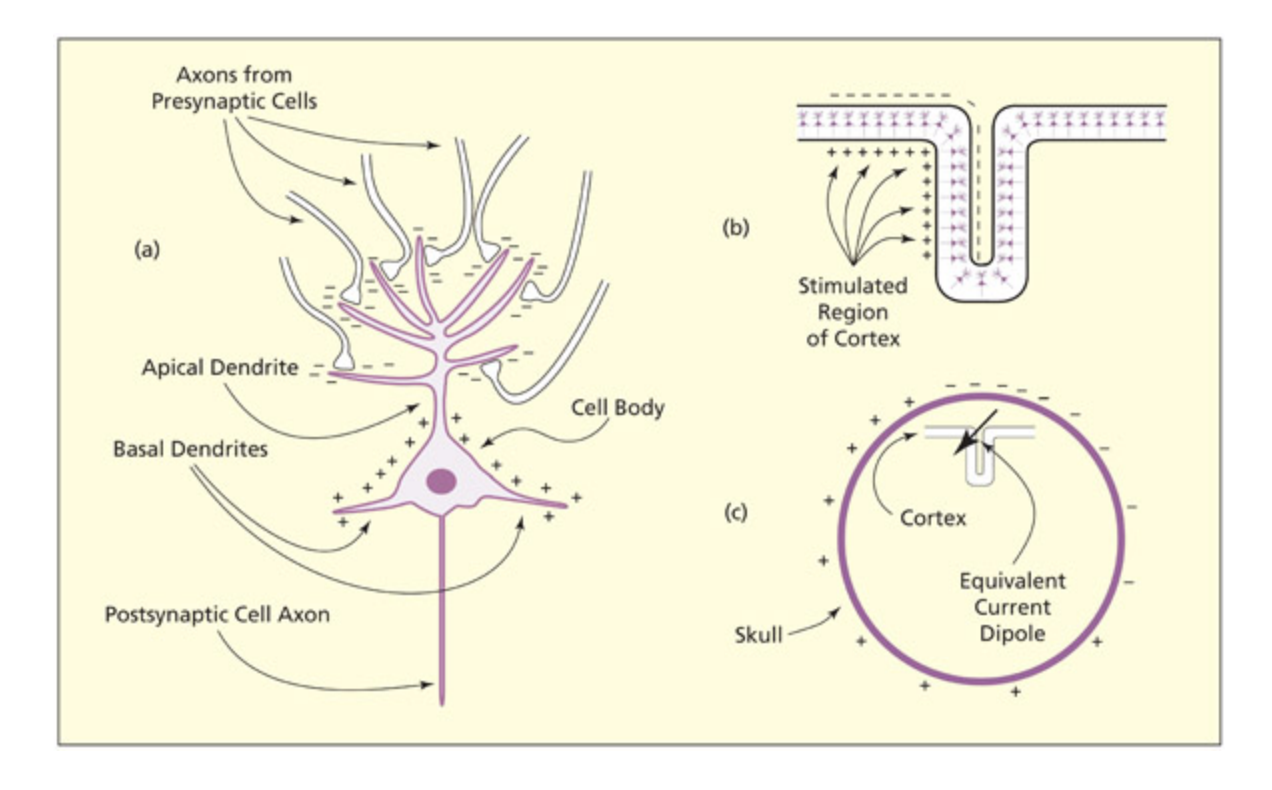
(a) Release of an excitatory neurotransmier results in positively arged ions flowing into the postsynaptic neuron (and a net negativity in the extracellular region). (b) is sets up a dipole that may sum together with dipoles from surrounding neurons (whi tend to be aligned in the same way). (c) is conducts to the scalp as a distribution of positive and negative arges. Changes in the negative or positive potential at a given site over time are the neural basis for the ERP signal. From Lu & Girelli, 1998.
📊 그림 설명
ERP 신호의 물리적 기초를 3단계로 설명하는 도해이다. (a) 흥분성 신경전달물질에 의해 양이온이 시냅스 후 뉴런으로 유입되어 세포 외 공간에 음성 전압이 형성되고, (b) 이 전기적 쌍극자(다이폴)가 병렬 배열된 주변 뉴런들과 합산되며, (c) 합산된 신호가 두피까지 전도되어 양성 및 음성 전하의 분포로 나타난다.
Whether a peak is positive or negative (its polarity) has no real significance in cognitive terms, nor does a positive peak reflect excitation and a negative peak inhibition. e polarity depends on the spatial arrangement of the neurons that are giving rise to the signal at that particular moment in time. Positive ions flow into the dendrites when an excitatory neurotransmier is released leaving a net negative voltage in the extracellular space. is creates what is called a dipole. Dipoles from different neurons and different regions summate and conduct to the skull, and these give rise to the aracteristic peaks and troughs of the ERP waveform. What is of interest in the ERP waveform, in terms of linking it to cognition, is the timing and amplitude of those peaks. is is considered in the next section.
NOTE
신호 catch가 가능한 건, 뉴런들이 일정하게 방향성을 가지고 정렬되어 있고, 서로 동조 즉, 비슷한 행동을 하는 걸 보이기 때문.
Key Terms
Dipole
A pair of positive and negative electrical charges separated by a small distance.
다이폴(dipole)은 양성과 음성의 전하가 짧은 거리로 분리된 전기적 쌍으로, 뉴런의 시냅스 후 전류에 의해 생성된다. 여러 뉴런의 다이폴이 합산되어 두피까지 전도되며, 이것이 ERP 파형의 물리적 기초를 형성한다.
시험 팁
ERP 정점 명명법을 정확히 이해하자. P300 = 자극 후 300ms에 나타나는 양성(Positive) 피크이지, 300번째 양성 피크가 아니다. N400도 마찬가지로 400ms의 음성(Negative) 피크이다. 정점의 극성(양성/음성)은 흥분/억제와 무관하며, 다이폴의 공간적 배열에 따라 결정된다는 점이 중요하다.
Some Practical Issues to Consider When Conducting EEG/ERP Research
Summary
**Picton et al. (2000)**이 11개 주요 연구실의 합의로 제시한 ERP 실험 지침은 실험 설계, 데이터 수집, 분석, 결과 해석의 체계적 기준을 제공한다. 이 지침은 연구의 재현성과 신뢰성을 높이기 위해 신규 연구자에게 필수적인 참고 자료로 권장된다.
Where can a set of guidelines for conducting and reporting ERP experiments be found?
A detailed set of guidelines is provided by Picton et al. (2000) and is based on a consensus agreed by 11 leading laboratories in the field. is is recommended reading for all new researers in the field.
What behavioral measures should be obtained?
Summary
ERP 실험에서는 대부분 외현적 행동 반응(버튼 누름)을 통해 반응 시간과 오류율을 분석하며, 구두 응답은 턱 움직임이 EEG 신호를 교란하므로 사용할 수 없다. 초기 가설은 특정 ERP 성분(예: P300의 지연 시간 변화)에 대한 구체적 예측을 명시해야 하며, 비구체적 예측은 대규모 데이터셋에서 허위 결과 발생 위험이 높다.
In almost all ERP experiments, participants are required to perform a task in whi an overt behavioral response is required (e.g. a buon press), and this can be analyzed independently (e.g. in terms of reaction times and/or error rates). One exception to this is ERP responses to unaended stimuli (e.g. ignored stimuli, stimuli presented subliminally). It is not possible to record vocal responses (e.g. picture naming) because jaw movements disrupt the EEG signal.
It is important that the initial hypothesis places constraints on the ERP component of interest (e.g. “the experimental manipulation will affect the latency of P300 component”) rather than predicting nonspecific ERP anges (e.g. “the experimental manipulation will affect the ERP in some way”). is is because the dataset generated from a typical ERP experiment is large and the ance of finding a “significant” result that is not justified by theory or reliable on replication is high.
How can interference from eye movement be avoided?
Summary
눈 움직임과 눈꺼풀 움직임은 EEG 신호와 동일한 주파수대에서 발생하는 주요 간섭 요인이다. 참가자에게 깜빡임을 억제하도록 지시하는 방법은 부차적 과제로 작용할 수 있고, 눈 움직임이 발생한 시행을 필터링하거나 제거하는 방법이 대안으로 사용된다.
Not all of the electrical activity measured at the scalp reflects neural processes. One major source of interference comes from movement of the eyes and eyelids. ese movements occur at the same frequencies as important components in the EEG signal. ere are a number of
ways of reducing or eliminating these effects. One can instruct the participant not to blink or to blink only at specified times in the experiment (e.g. aer making their response). e problem with this method is that it imposes a secondary task on the participant (the task of not moving their eyes) that may affect the main task of interest. It is also possible to discard or filter out the effect of eye movements in trials in whi they have occurred (Lu, 2005).
Mental Chronometry in Electrophysiology and Cognitive Psychology
Summary
정신적 시간 측정(mental chronometry)은 반응 시간과 ERP를 통해 인지 프로세스의 시간적 특성을 탐구하는 분야이다. 정보 처리의 효율성이나 본질의 변화가 과제 수행 시간에 반영된다는 기본 원리에 기반하며, EEG/ERP를 활용하여 인지 단계의 타이밍 구조를 분석한다.
Key Terms
Mental chronometry
The study of the time course of information processing in the human nervous system.
정신적 시간 측정(mental chronometry)은 인간 신경계에서 정보 처리의 시간 경로를 연구하는 분야이다. 예를 들어, 4+2=6의 검증이 5+3=8보다 빠르다는 사실은 수학적 합이 단순 사실로 저장되지 않으며, 수치 크기가 처리 효율에 영향을 미침을 시사한다. 반응 시간의 상대적 차이를 통해 인지 프로세스의 본질을 추론할 수 있다.
Mental ronometry can be defined as the study of the time-course of information processing in the human nervous system (Posner, 1978). e basic idea is that anges in the nature or efficiency of information processing will manifest themselves in the time it takes to complete a task. For example, participants are faster at verifying that 4 + 2 = 6 than they are to verify that 4 + 3 = 7, and this is faster than verifying that 5 + 3 = 8 (Parkman & Groen, 1971). What can be concluded from this? First of all, it suggests that mathematical sums su as these are not just stored as a set of
facts. If this were so, then all the reaction times would be expected to be the same because all statements are equally true. It suggests, instead, that the task involves a stage in processing that encodes numerical size together with the further assumption that larger sums place more limits on the efficiency of information processing (manifested as a slower verification time). is provides one example of how it is possible to make inferences about the nature of cognitive processes from timing measures.
Key Terms
Additive factors method
A general method for dividing reaction times into different stages.
추가 요인 방법(additive factors method, Sternberg 1969)은 반응 시간을 인코딩, 비교, 결정, 반응의 단계로 분리하는 방법이다. 서로 다른 단계에 영향을 주는 변수는 가산적 효과를, 동일 단계에 영향을 주는 변수는 상호작용적 효과를 나타낸다. ERP 분석에서도 각 정점이 인지 단계와 대응될 수 있으나, 여러 신경 집단의 활동이 중첩되어 일대일 대응은 어렵다.
Additive factors method
A task su as verification of sums is likely to involve a series of stages, including visual recognition of the digits, computing the sum and producing a response. e reaction time measure is the end product of all these stages. Sternberg (1969) developed a general method for dividing reaction times into different stages termed the additive factors method. His experiment involved a working memory task in whi participants were given an array of one, two or four digits to hold in mind (e.g. 5, 9, 3, 2). ey were then shown a single probe digit (e.g. 9) and asked to press one of two buons (labeled “yes” and “no”) to indicate whether this item had been in the previous array. Sternberg proposed that the task could be divided into a number of separate stages, including:
-
- Encoding the probe digit.
-
- Comparing the probe digit with the items held in memory.
-
- Decision about whi response to make.
-
- Responding by executing the buon press.
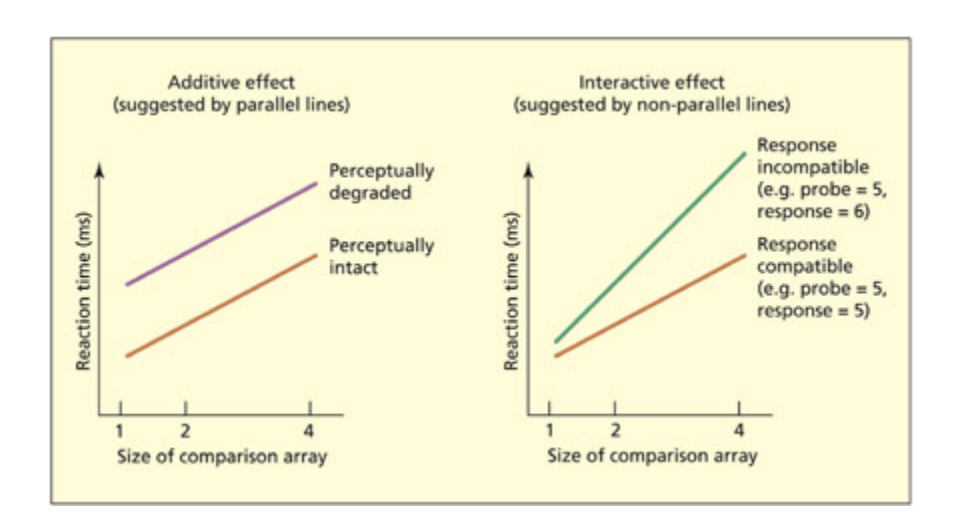
Sternberg’s additive factors method assumes that if two variables affect different stages of processing then they should have an additive effect on the overall reaction time (le), but if two variables affect the same stage of processing then the factors should have an interactive effect (right). His task involved comparing a probe digit (e.g. 5) with an array of one, two, or four digits held in mind.
📊 그림 설명
Sternberg의 추가 요인 방법을 도식화한 그림이다. 두 변수가 서로 다른 처리 단계에 영향을 미치면 반응 시간에 가산적 효과(좌)를, 동일 단계에 영향을 미치면 상호작용적 효과(우)를 나타냄을 보여준다. 탐사 숫자를 1, 2, 4개의 기억 항목과 비교하는 작업 기억 과제가 예시로 사용된다.
실험 상세 — Sternberg (1969): 추가 요인 방법 (Additive Factors Method)
목적: 반응 시간이라는 단일 측정치를 여러 개의 독립적인 인지 처리 단계로 분리할 수 있는 일반적 방법론을 개발한다.
방법: 참가자에게 1개, 2개, 또는 4개의 숫자 배열(예: 5, 9, 3, 2)을 기억하게 한 뒤, 단일 탐사 숫자(probe digit, 예: 9)를 제시하고 “Yes/No” 버튼을 눌러 해당 숫자가 배열에 있었는지 판단하게 했다.
핵심 논리 — 과제의 4단계 분리:
- 인코딩 (Encoding) — 탐사 숫자를 시각적으로 인식
- 비교 (Comparison) — 탐사 숫자를 기억 속 항목들과 대조
- 결정 (Decision) — 어떤 반응을 할지 판단
- 반응 (Response) — 버튼 누르기 실행
가산적 효과 vs 상호작용적 효과:
- 두 변수가 서로 다른 단계에 영향 → 전체 반응 시간에 가산적(additive) 효과 (각 변수의 효과가 독립적으로 합산)
- 두 변수가 같은 단계에 영향 → 상호작용적(interactive) 효과 (한 변수의 효과가 다른 변수의 수준에 따라 달라짐)
예시:
- 탐사 숫자의 지각적 선명도(예: 패턴 배경 위에 제시) → 인코딩 단계에 영향
- 기억 배열의 항목 수(1개 vs 4개) → 비교 단계에 영향
- 이 두 변수가 가산적 효과를 보이면 → 서로 다른 단계에 영향을 미친다는 증거
의의:
- 미지의 변수(예: 수면 부족, 파킨슨병, 읽기 능력)가 어떤 인지 단계에 영향을 미치는지 실험적으로 판별할 수 있는 범용 도구를 제공
- 이후 ERP 연구에서 각 피크/트로프를 인지 단계와 대응시키는 접근법의 이론적 토대가 됨
- 한계: 각 단계가 엄격하게 순차적(이전 단계가 끝나야 다음 단계 시작)이라고 가정하나, 실제로는 단계들이 중첩될 수 있어 이 가정이 항상 성립하지는 않음
He further postulated that ea of these stages could be independently influenced by different factors affecting the task. For instance, the encoding stage may be affected by the perceptibility of the probe digit (e.g. presenting it on a paerned baground). e comparison stage may be affected by the number of items in the array (the more items in the array, the slower the task). He reasoned that, if different factors affect different stages of processing, then the effects should have additive effects on the overall reaction time, whereas if they affect the same processing stage, they should have interactive effects. e strength of this method is that one could then take an unknown factor (e.g. sleep deprivation, Parkinson’s disease, reading ability) and determine whether this has an interactive effect on stimulus perceptibility (implying that the new factor affects perceptual encoding) or whether it has an interactive effect with the number of items in the array (implying the new factor affects the comparison stage) or both (implying the new factor has effects at multiple levels).
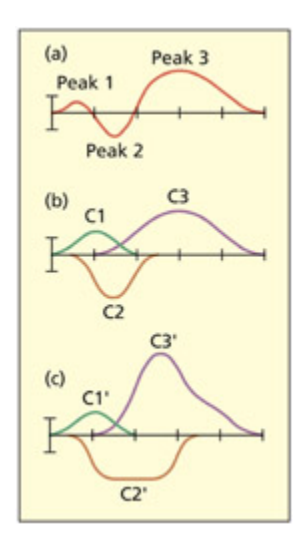
Graph (a) shows an observed ERP waveform and graphs (b) and (c) show two different sets of hidden components that could have given rise to it. is illustrates the point that there is not a one-to-one mapping between ERP components and the activity of underlying cognitive/neural components. From Lu, 2005. © 2004 Massauses Institute of Tenology by permission of the MIT Press.
📊 그림 설명
관찰된 ERP 파형(a)과 이를 생성할 수 있는 두 가지 서로 다른 숨겨진 성분 조합(b, c)을 보여준다. 동일한 ERP 파형이 다양한 인지/신경 성분의 조합에서 생성될 수 있으므로, ERP 성분과 기저 인지 프로세스 간에 일대일 대응이 성립하지 않음을 시각적으로 설명한다.
e additive factors approa has been very influential in cognitive psyology resear, although it is to be noted that the assumptions do not always apply. For example, the model assumes that the stages are strictly sequential (i.e. later stages do not occur until earlier ones are complete), but this assumption is not always valid.
At this juncture it is useful to consider how the mental ronometry approa applies to the analysis and interpretation of ERP data. Whereas a reaction time consists of a single measure that is assumed to reflect different stages/components, an ERP waveform consists of a series of peaks and troughs that vary continuously over time. ese peaks and troughs are likely to have some degree of corre spondence with different cognitive stages of processing. For example, in the task described above, earlier peaks may reflect perceptual encoding and later peaks may reflect the comparison stage. One could then observe how the amplitude of those peaks varied, say, with the number of items to be compared. One could also observe whether a new variable (e.g. sleep deprivation) affected earlier or later peaks. e different peaks and troughs of the ERP signal have been referred to as ERP components (Donin, 1981). ere may not be a simple mapping between
an ERP component and a cognitive component of a task. For example, a single cognitive component may reflect the action of several spatially separate neural populations (i.e. one cognitive component could affect several ERP components) or several cognitive components may be active at once and sum together, or cancel ea other out, in the ERP waveform (i.e. several cognitive components affect a single ERP component). As su, some researers prefer to use the more neutral term ERP deflection rather than ERP component.
Investigating face processing with ERPs and reaction times
Summary
N170(170ms 음성 피크)은 얼굴의 지각적 인코딩 단계를 반영하며 유명성과 무관하지만 지각적 퇴행 시 감소한다. N250은 유명하거나 익숙한 얼굴에 더 크게 반응하여 구체적 얼굴 특성의 인코딩과 관련된다. 300ms 이후 양성 성분은 인물 정체성의 반복과 익숙성에 민감하며 이름에도 일반화되어, 얼굴 인식이 모달리티에 구애받지 않는 개인 정체성 표현을 포함함을 시사한다.
is apter has already considered the neural representation of faces as measured by single-cell recordings. ERP studies have also investigated the way that faces are processed. A full model of face processing is discussed in Chapter 6, but a consideration of a few basic stages will suffice for the present needs. An initial stage consists of perceptual coding of the facial image (e.g. location of eyes, mouth), followed by a stage in whi the facial identity is computed. is stage is assumed to map the perceptual code onto a store of known faces and represents the face irrespective of viewing conditions (e.g. lighting, viewing angle). (Note that this doesn’t assume grandmother cells because facial identity could be computed by a population of neurons.) Finally, there may be a representation of the identity of the person that is not tied to any modality (e.g. responds to faces and names) and may enable retrieval of other types of knowledge (e.g. their occupation).
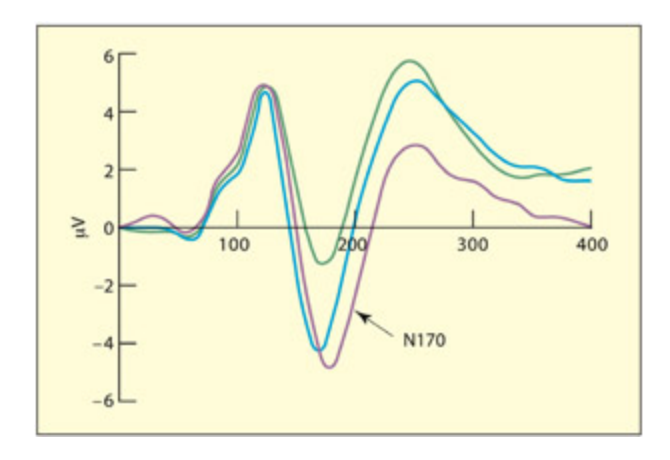
As with the single-cell results, there is evidence for an ERP component that is relatively selective to the processing of faces compared with other classes of visual objects. is has been termed the N170 (a negative peak at 170 ms) and is strongest over right posterior temporal electrode sites (Bentin et al., 1996). is component is uninfluenced by whether the face is famous or not (Bentin & Deouell, 2000) and is also found for cartoon “smiley” faces (Sagiv & Bentin, 2001). It is, however, reduced if the face is perceptually degraded (Sweinberger, 1996). e N250, by contrast, is larger for famous
and personally familiar faces relative to unfamiliar faces (Herzmann, et al., 2004) and responds to presentation of different images of the same person (Sweinberger et al., 2002b). is suggests that it codes properties of the specific face rather than the specific image. Later, positive-going com ponents (from 300 ms onwards) are also sensitive to the repetition and familiarity of specific person identities, and the effects generalize to names as well as faces (Sweinberger et al., 2002a).
Key Terms
N170
An ERP component (negative potential at 170 ms) linked to perceiving facial structure.
N170은 얼굴 구조 인식과 연관된 ERP 성분으로, 인간 및 동물 얼굴에서 관찰되지만 다른 물체에서는 나타나지 않는다. 연관성 프라이밍(associative priming)은 ERP 분석 결과 300ms 이후에 영향을 미쳐 지각 후 처리 단계와 관련되며, 이는 Sternberg의 이산 단계 가정이 항상 유효하지 않음을 시사한다.
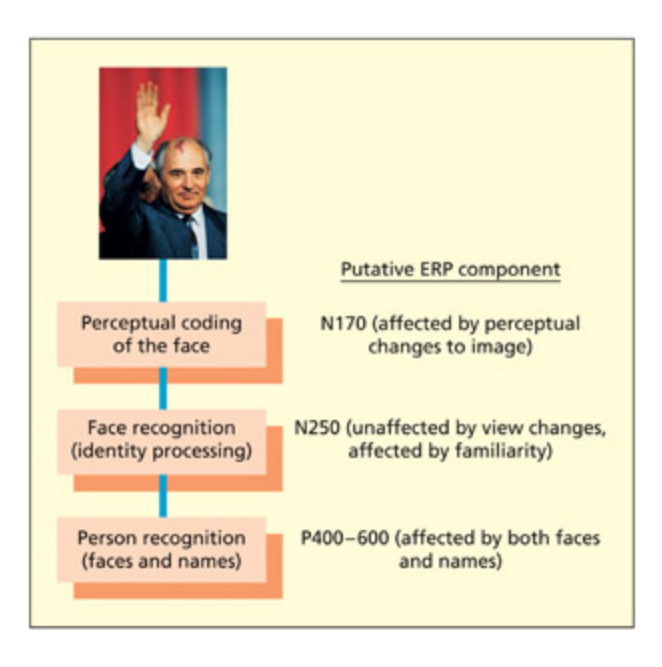
A simple model of several hypothetical stages involved in face processing together with their putative ERP manifestations.
📊 그림 설명
얼굴 처리에 관여하는 여러 가설적 단계(지각적 인코딩, 얼굴 정체성 계산, 개인 정체성 표현)와 각 단계에 대응하는 ERP 성분(N170, N250, 300ms 이후 양성 성분)의 관계를 도식화한 모델이다.
Photo © Bernard Bisson and ierry Orban/Sygma/Corbis.
e N170 is observed for both human faces (purple) and animal faces (blue), but not other objects (green).
📊 그림 설명
N170 ERP 성분이 인간 얼굴(보라색)과 동물 얼굴(파란색)에서는 관찰되지만, 다른 물체(녹색)에서는 나타나지 않음을 보여주는 파형 비교 그래프이다. N170이 얼굴 범주에 선택적으로 반응하는 지각적 성분임을 실증한다.
From Rousselet et al., 2004. With permission of ARVO.
Having sketed out a plausible relationship between different components of the ERP wave form and different cognitive processes, it is possible to use these electrophysiological markers to adjudicate between different theories of face processing. One debate in the cognitive psyology literature concerns the locus of associative priming. Associative priming refers to the fact that reaction times are faster to a stimulus if that stimulus is preceded by a stimulus that tends to co-occur with it in the environment. For example, judging that the face of Mikhail Gorbaev (the last President of the Soviet Union) is familiar is performed faster if it immediately follows Boris Yeltsin’s face (former President of Russia) or even Yeltsin’s name (Young et al., 1988). e fact that associative priming is found between names and faces might imply that the effect arises at a late stage of processing. However, there is evidence inconsistent with this. Using Sternberg’s (1969) method, it has been found that associative priming interacts with stimulus degradation (Bruce & Valentine, 1986) and that associative priming interacts with how perceptually distinctive a face is (Rhodes & Tremewan, 1993). is would imply that associative priming has a perceptual locus su that perceiving Gorbaev’s face also activates the
perceptual face representation of Yeltsin. Sweinberger (1996) used ERP measures to determine the locus of associative priming of faces and names. ERP was suitable for addressing this question because it enables early and late time points to be measured separately. He found that associative priming has a late effect (aer 300 ms) on the ERP waveform that is more consistent with a post-perceptual locus. Effects of stimulus degradation were found under 150 ms. Swein-berger (1996) suggests that, in this instance, the Sternberg method may have led to an invalid conclusion because it assumes discrete stages.
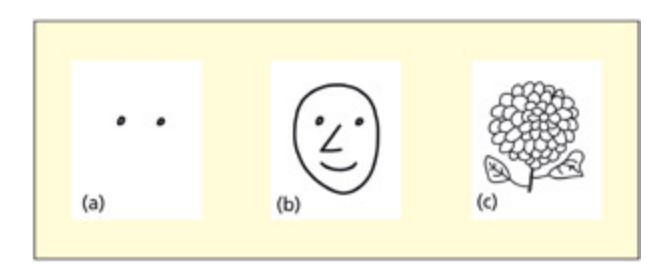
Two horizontally spaced symbols (the dots in a) do not elicit an N170 unless they have previously been presented in the context of a face (b). e participant’s task was merely to count flowers (e.g. c), and so both the faces and “eyes” were irrelevant to the task.
📊 그림 설명
수평으로 배치된 두 점(a)은 단독으로는 N170을 유발하지 않지만, 이전에 얼굴 맥락에서 눈으로 제시된 경우(b)에는 N170이 나타남을 보여준다. 참가자의 과제는 꽃 세기(c)였으므로 얼굴과 “눈” 자극은 과제와 무관했으며, 이는 N170이 순수한 외생성 성분이 아닌 맥락 의존적 성분임을 시사한다.
From Bentin et al., 2002. Reprinted by permission of Blawell Publishing.
Endogenous and exogenous ERP components
Summary
ERP 성분은 자극의 물리적 특성에 의존하는 외생성(exogenous)과 과제 특성에 의존하는 내생성(endogenous)으로 분류되며, 외생성이 일반적으로 먼저 나타난다. 그러나 이 구분은 범주적이라기보다 연속적 차원에 가까우며, 예를 들어 N170은 지각적 성분이지만 이전의 얼굴 맥락에 따라 단순 기호에서도 유발될 수 있다.
Traditionally, ERP components have been classified as belonging to one of two categories. Exogenous components are those that appear to depend on the physical properties of a stimulus (e.g. sensory modality, size, intensity). ese have also been called evoked potentials. Endogenous components, in contrast, appear to depend on properties of the task (e.g. what the participant is required to do with the stimulus). ese can even occur in the absence of an external stimulus (e.g. if an expected stimulus does not occur; Suon et al., 1967). Exogenous components tend to be earlier than endogenous components.
Although the exogenous–endogenous classification is useful, it should be considered as a dimension rather than a true categorical distinction. To remain with the current example of face processing, consider the nature of the ERP waveform when viewing two repeated symbols that are horizontally spaced (e.g. + +). Typically, su symbols do not evoke the N170 response aracteristic of face processing (Bentin et al., 2002). However, if the symbols have previously been shown embedded in a face context (as eyes), then the pair of symbols do elicit the N170 response (Bentin et al., 2002). Is this an endogenous or exogenous component? It is impossible to say. Although the N170 is normally taken as indicative of perceptual processing (an exogenous component), in this instance it is entirely dependent on the interpretive bias given.
Key Terms
Associative priming
Reaction times are faster to stimulus X after being presented to stimulus Y if X and Y have previously been associated together (e.g. if they tend to co-occur).
연관성 프라이밍(associative priming)은 자극 X와 Y가 연관되어 있을 때, Y 제시 후 X에 대한 반응 시간이 빨라지는 현상이다. ERP 연구에서 이 효과는 300ms 이후에 나타나 지각 후 처리 단계와 관련됨이 확인되었다.
Exogenous
Related to properties of the stimulus.
외생성(exogenous) ERP 성분은 자극의 물리적 특성(감각 모드, 크기, 강도)에 의존하며 초발성 전위(evoked potentials)로도 불린다. P100, N170 등이 대표적이며, 인지적 처리와 무관하게 자극의 물리적 속성에 반응한다.
Endogenous
Related to properties of the task.
내생성(endogenous) ERP 성분은 과제의 인지적 요구(주의 집중, 결정 과정)에 의존하며, 외부 자극 없이도 발생할 수 있다. P300이 대표적 예로, 자극의 물리적 특성이 아닌 인지적 작업의 복잡성과 연관된다.
Inverse problem
The difficulty of locating the sources of electrical activity from measurements taken at the scalp (in ERP research).
역 문제(inverse problem)는 두피에 측정된 전기 신호로부터 뇌 내 전기 활동의 원천 위치를 추정하는 어려움을 의미한다. 동일한 두피 신호가 다양한 뉴런 활동 패턴에서 유도될 수 있어 수학적으로 무한한 해가 존재하며, 이는 ERP의 공간 해상도가 낮은 근본적 원인이다.
임상 사례
N170 성분은 얼굴 인식 연구에서 핵심적이다. 자폐 스펙트럼 장애(ASD) 환자에서 N170이 지연되거나 감소한다는 연구 결과가 있으며, 이는 얼굴 지각의 초기 처리 단계에 이미 차이가 있음을 시사한다. 외생성(exogenous) vs 내생성(endogenous) 구분은 시험에서 자주 묻는 개념이므로, “외생성 = 자극 특성, 내생성 = 과제 특성”으로 구분하자.
Dipole modeling
Summary
다이폴 모델링(dipole modeling)은 역 문제를 해결하기 위해 신호 생성에 기여하는 다이폴의 수를 가정하여 원천 위치를 추정하는 방법이다. 그러나 가정에 따라 결과가 달라질 수 있어 정확한 해를 보장하지 못하며, 보다 정밀한 공간 해상도를 위해 fMRI나 MEG가 보완적으로 활용된다.
An aempt to solve the inverse problem in ERP resear that involves assuming how many dipoles (regions of electrical activity) contribute to the signal recorded at the scalp.
The spatial resolution of ERPs
Summary
ERP의 공간 해상도가 낮은 이유는 역 문제(inverse problem) 때문으로, 두피 신호의 원천 위치와 강도에 대해 수학적으로 무한한 해가 존재한다. 다이폴 모델링으로 N170과 N250의 원천을 각각 후두부와 방추형 회(fusiform gyrus)로 추정하였으나 정확도에 한계가 있다. 보다 정밀한 공간 해상도를 위해 fMRI나 MEG와의 다중 모드 접근이 필요하다.
e discussion so far has emphasized the importance of ERPs in the timing of cognition. e reason why the spatial resolution of this method is poor is given by the so-called inverse problem. If one had, say, three sources of electrical activity in the brain during a given task, and the magnitude and location of the activity were known, then it would be possible to calculate the electrical potential that we would expect to observe some distance away at the scalp. However, this is not the situation that is encountered in an ERP study; it is the inverse. In an ERP study, the electrical potential at the scalp is known (because it is measured), but the number, location, and magnitude of the electrical sources in the brain are unknown. Mathematically, there are an infinite number of solutions to the problem.
e most common way of aempting to solve the inverse problem involves a procedure called dipole modeling. is requires assumptions to
be made about how many regions of the brain are critical for generating the observed paern of scalp potentials. Aempts at dipole modeling with the N250 and N170 evoked by face processing (see above) revealed probable loci in the fusiform gyrus and the posterior occipital region, respectively (Sweinberger et al., 2002b). However, the most common way of obtaining good spatial resolution is to use a different method altogether, su as fMRI (see Chapter 4) or magnetoencephalography (MEG). (For similar results from fMRI concerning face processing, see Eger et al., 2004.)
Why are Caricatures Easy to Recognize?
Summary
카리커처는 개인의 독특한 특징을 과장하여 실제 얼굴보다 최대 2배 빠르게 인식될 수 있다. 기억 표현 자체가 카리커처화되어 있다는 가설과, 지각적 독특성이 유사 얼굴과의 경쟁을 줄여 인식을 돕는다는 가설이 있다. ERP 연구에서 카리커처가 N170 성분을 증가시키는 결과는 후자의 가설, 즉 지각적 독특성이 인식 효율의 주요 원인임을 지지한다.

is caricature is instantly recognizable despite significant distortions. We are sometimes faster at recognizing caricatures than actual depictions. Why might this be?
📊 그림 설명
상당한 왜곡에도 불구하고 즉시 알아볼 수 있는 카리커처의 예시이다. 개인의 독특한 특징이 과장되어 있으며, 실제 얼굴보다 카리커처를 더 빠르게 인식하는 현상의 원인을 탐구하는 맥락에서 제시된다.
Caricatures of faces are typically considered humorous and are oen used for deliberate moery or propaganda. As Riard Nixon’s unpopularity grew during the Watergate scandal, so did his nose and jowls in published caricatures (see Rhodes, 1996). e paradox of caricatures is that the face is instantly recognizable despite being perceptibly wrong. In fact, people can sometimes be twice as fast at
recognizing a caricature of a face as the same face undistorted (Rhodes et al., 1987); the caricature appears to be more like the face than the face itself. What does this reveal about the way that faces are processed and represented?
First of all, it is important to clarify how caricatures are created. Caricatures exaggerate the distinctive features of an individual. Computer routines now exist that compare, for example, the size of an individual’s nose with the average nose size. If the person has a larger than average nose, then this will be enlarged further in the caricature. If someone has a smaller than average nose, it will be shrunk in the caricature. It is also possible to morph a face to make it look more average (a so-called anti-caricature), and su faces are typically rated as more aractive than the real or caricatured face. One explanation for the effect of caricatures is to assume that our memory representations of faces are caricatured themselves; that is, we store the distinctive properties of a face rather than the face as it is. However, explanations su as these must assume that a “norm” or prototype face exists from whi to infer what constitutes a distinctive feature. Another hypothesis is that it is the distinctiveness of caricatures per se that aids their recognition because there are fewer similar-looking competitor faces (Valentine, 1991). is account does not need to assume the existence of a face prototype, or that the stored representations themselves are caricatured. Resear using ERPs is consistent with this view. Photographic caricatures of unfamiliar people lead, initially, to an enhancement of the N170 component relative to undistorted images or anti-caricatures (Kaufmann & Sweinberger, 2008). As this component is normally associated with perceptual coding of faces rather than memory of faces, it suggests that the effect is more likely to be due to perceptual distinctiveness than the way faces are coded in memory.
Evaluation
Investigating the time-course of cognitive processes is an important method in cognitive psyology and cognitive neuroscience. Event-related potentials have excellent temporal resolution. is method has a number of benefits over and above reaction-time measurements: it provides a continuous measurement of anges over time (rather than a single timing measure) and it is, at least in theory, easier to link to neural processes in the brain. ERP also enables electrophysiological anges associated with unaended stimuli (that are not responded to) to be measured whereas a reaction-time measure always requires an overt behavioral response.
Magnetoencephalography (MEG)
Summary
MEG는 뇌의 자기장을 SQUID 센서로 측정하며, 신호가 두피와 뇌막의 영향을 받지 않아 2-3mm 수준의 공간 해상도를 제공한다. EEG보다 공간 해상도가 우수하지만 액체 헬륨 냉각과 자기장 차폐실이 필요하여 고비용이다. MEG는 뇌 고랑(sulci) 활동에 민감하고 깊은 다이폴 감지에는 약하며, EEG와 상호 보완적으로 활용된다.
e recording of magnetic signals, as opposed to electrical ones, generated by the brain has a mu shorter history in cognitive neuroscience and still remains in its infancy (for reviews, see Papanicolaou, 1995; Singh, 2006). All electric currents, including those generated by the brain, have an associated magnetic field that is potentially measurable. However, the size of this field is very small relative to the ambient magnetic field of the earth. As su, the development of magneto encephalography (MEG) had to wait for suitable tenological advances to become a viable enterprise. is tenological advance came in the form of superconducting devices termed SQUIDs (an acronym of Superconducting antum Interference Device). A whole-head MEG contains 200–300 of these devices. e apparatus used requires extreme cooling, using liquid helium, and isolation of the system in a magnetically shielded room. As su, the costs and practicalities associated with MEG are far greater than those needed for EEG. However, the biggest potential advantage of MEG over EEG is that it permits a mu beer spatial resolution.
An MEG scanner. is extremely powerful maine measures the magnetic fields produced by electrical activity in the brain.
📊 그림 설명
MEG 스캐너의 외관을 보여주는 사진이다. 이 장비는 SQUID 센서를 사용하여 뇌의 전기적 활동이 생성하는 미세한 자기장을 측정하며, 액체 헬륨으로 냉각되는 초전도 장치와 자기장 차폐실 내에서 운용된다.
| MEG | EEG/ERP |
|---|---|
| Signal unaffected by skull, meninges, etc. | Signal affected by skull, meninges, etc. |
| Poor at detecting deep dipoles | Detects deep and shallow dipoles |
| More sensitive to activity at sulci | Sensitive to gyri and sulci activity |
| Millisecond temporal resolution | Millisecond temporal resolution |
| Potentially good spatial resolution(2–3 mm) | Poor spatial resolution |
| Expensive and limited availability | Cheaper and widely available |
Key Terms
Magnetoencephalography (MEG)
A noninvasive method for recording magnetic fields generated by the brain at the scalp.
MEG는 뇌에서 생성되는 자기장을 비침습적으로 기록하는 기술로, EEG와 동일한 밀리초 단위 시간 해상도를 제공하면서 더 높은 공간 해상도를 갖는다. 두피의 전기적 간섭이 적어 신경 활동의 원천 위치 추정에 유리하며 역 문제 해결에 기여한다.
시험 팁
시간 해상도 vs 공간 해상도 비교는 단골 출제 주제이다. EEG/ERP = 시간 해상도 우수(ms 단위), 공간 해상도 낮음. fMRI = 공간 해상도 우수(mm 단위), 시간 해상도 낮음(초 단위). MEG = 시간 해상도 우수 + 공간 해상도도 EEG보다 좋음(2-3mm), 단 비용이 매우 높다. EEG의 공간 해상도가 낮은 근본 원인은 **역 문제(inverse problem)**이다.
Summary and Key Points of the Chapter
Summary
뉴런 활동은 전기장과 자기장을 생성하며, 단일 세포 기록(침습적)과 EEG(비침습적)로 측정된다. 뉴런 집단의 동기화된 활동을 자극 시작과 연관시켜 평균화하면 ERP가 생성되며, 자극이나 과제 조건을 변화시키면 ERP 파형의 특정 부분에 체계적 변화가 나타나 인지 프로세스의 시간적 순서와 독립성을 추론할 수 있다.
- Neuronal activity generates electrical and magnetic fields that can be measured either invasively (e.g. single-cell recording) or noninvasively (e.g. EEG).
- Studies of single-cell recordings are based on measuring the number of action potentials generated and provide clues about how neurons code information, by measuring the specificity of their responses to external stimuli.
- When populations of neurons are active in synrony they produce an electric field that can be detected at the scalp (EEG). When many su waves are averaged together and linked to the onset of a stimulus (or response), then an event-related potential (ERP) is obtained.
- An ERP waveform is an electrical signature of all the different cognitive components that contribute to the processing of that stimulus. Systematically varying certain aspects of the stimulus
or task may lead to systematic variations in particular aspects of the ERP waveform. is enables inferences to be drawn about the timing and independence of cognitive processes.
Example Essay Questions
Summary
뉴런은 레이트 코딩과 시간적 코딩으로 정보를 인코딩하며, 단일 세포 기록과 EEG/ERP로 측정된다. N170 등 ERP 성분은 얼굴 처리의 시간적 단계를 반영하고, 역 문제로 인한 공간 해상도 한계는 다이폴 모델링과 MEG로 보완된다. 인지 신경과학의 핵심 과제는 이러한 전기생리학적 기법을 통합하여 인지 프로세스의 시공간적 구조를 규명하는 것이다.
- How does the brain generate electrical signals, and how are these used in electrophysiological teniques?
- How do neurons code information?
- What is an “event-related potential” (or ERP) and how can it be used to inform theories of cognition?
- What have electrophysiological studies contributed to our understanding of how faces are represented and processed by the brain?
Recommended Further Reading
Summary
EEG/ERP 연구의 핵심 참고서로 **Luck(2005)**의 ERP 기법 입문서, **Dittrich & Kieffaber(2014)**의 ERPLAB 및 주파수 분석 교재, **Senior et al.(2006)**의 단일 세포 기록/EEG/MEG를 포괄하는 실험 방법론서가 추천된다.
- Diter, C. L. & Kieffaber, P. D. (2014). EEG methods for the psychological sciences. London: Sage. Covers some material missed by Lu, including use of ERPLAB and frequency-based analysis.
- Lu, S. J. (2005). An introduction to the event-related potential technique. Cambridge, MA: MIT Press. is is the place to start if you are going to conduct resear using EEG/ERPs.
- Senior, C., Russell, T., & Gazzaniga, M. S. (2006). Methods in mind. Cambridge, MA: MIT Press. Includes apters on singlecell recording, EEG, and MEG.